Modeling the Spatial Distribution of Three Typical Dominant Wetland Vegetation Species’ Response to the Hydrological Gradient in a Ramsar Wetland, Honghe National Nature Reserve, Northeast China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Methods
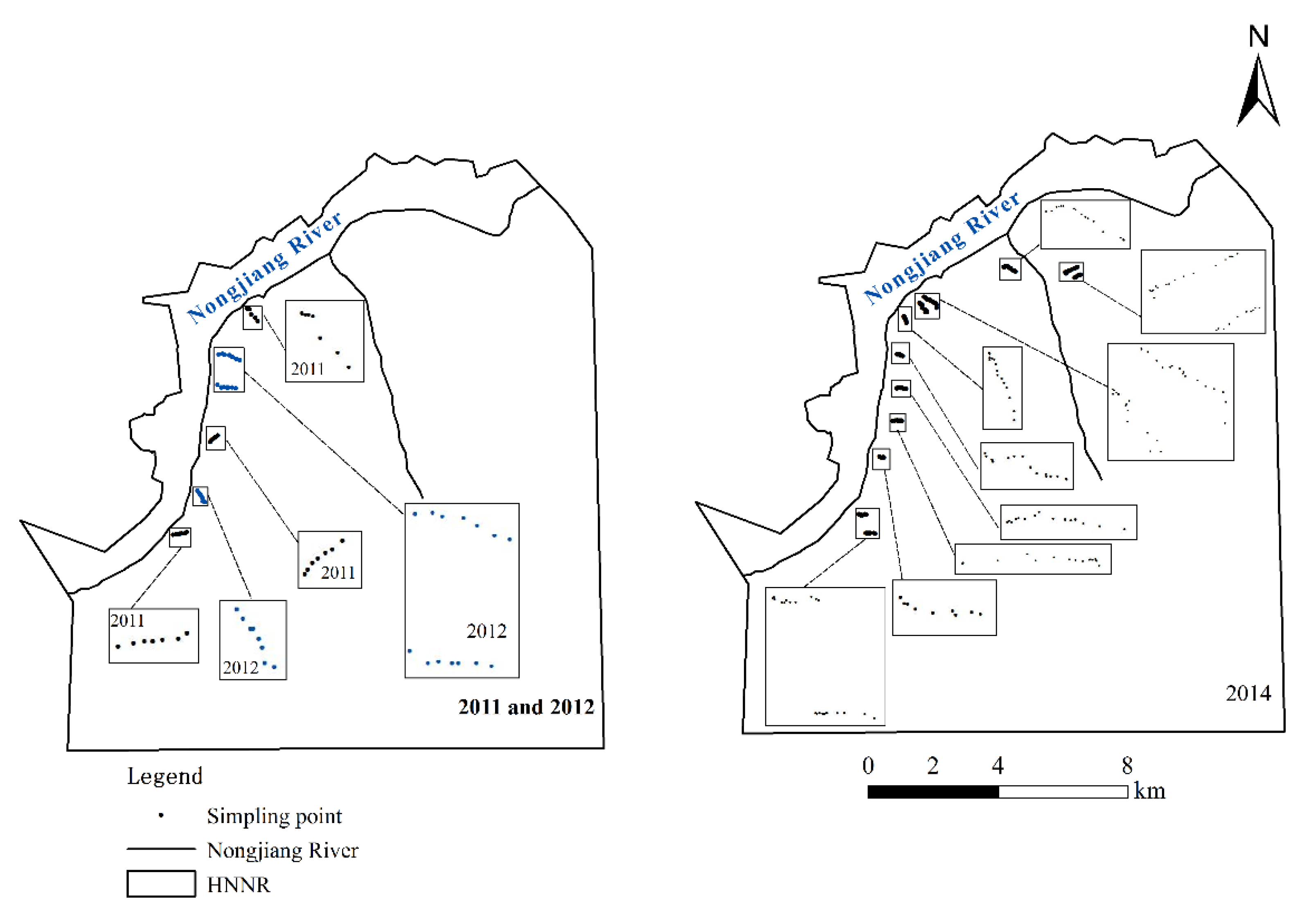
2.2.1. Sampling Method
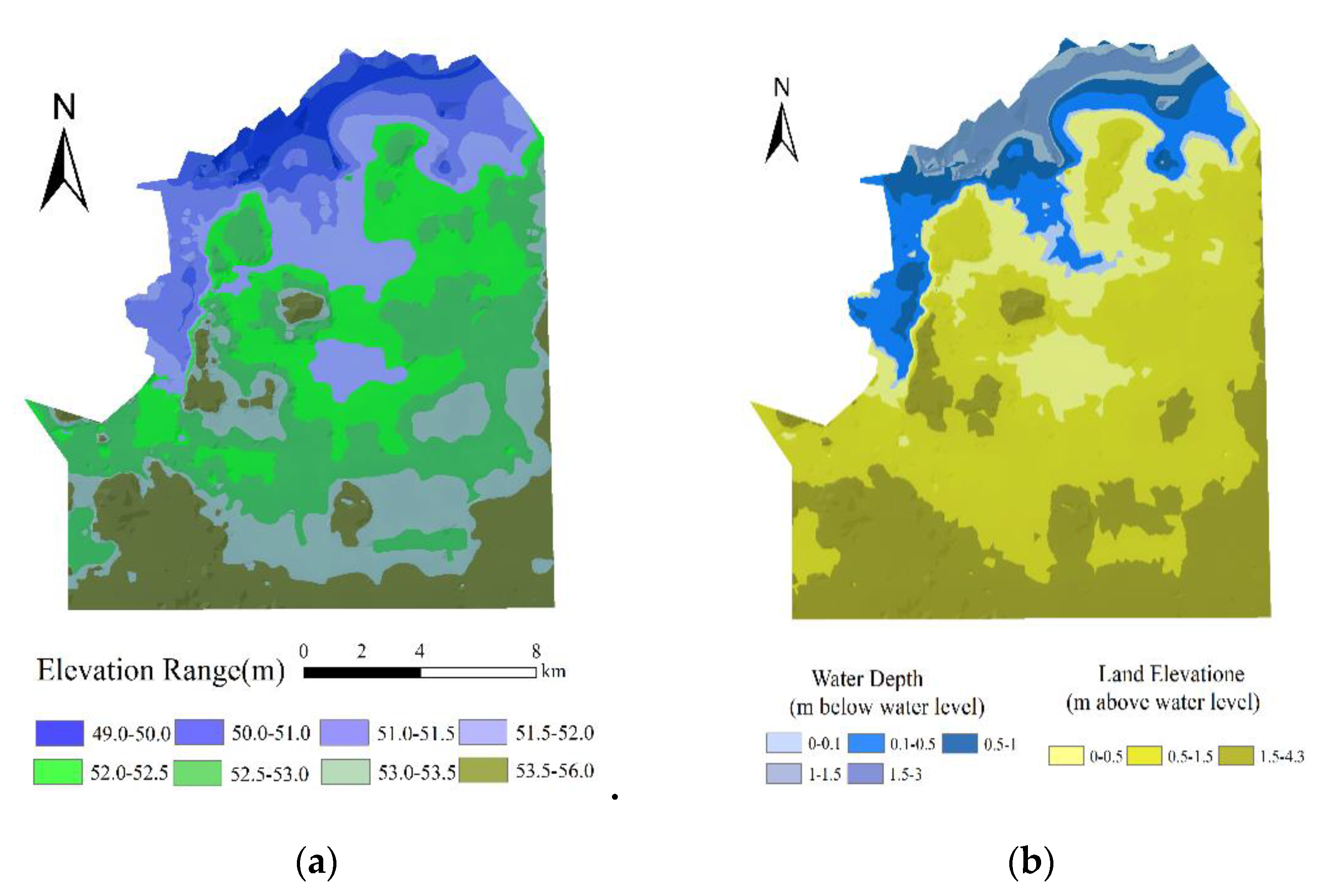
2.2.2. Hydrological Data
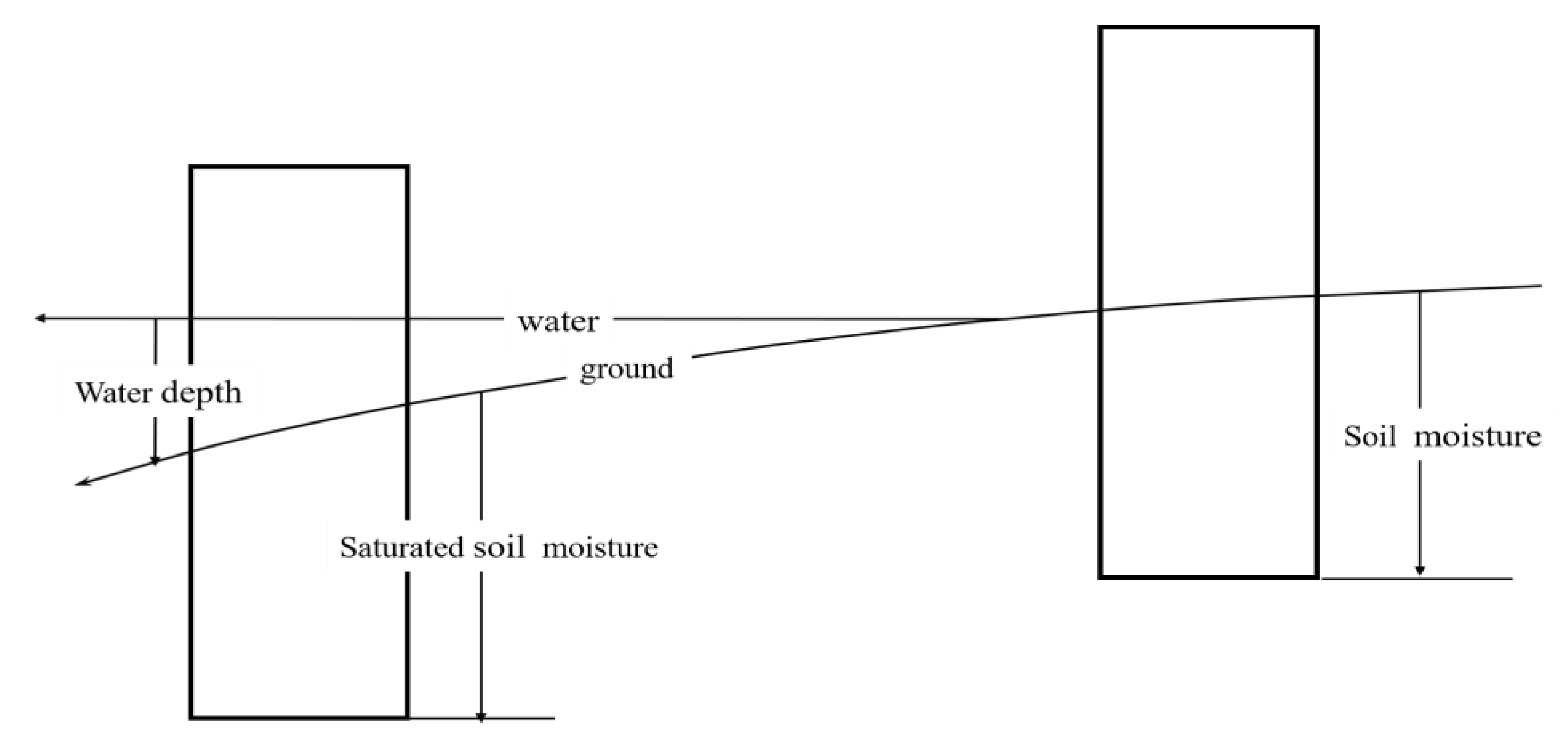
2.2.3. Conversion of the Hydrological Gradient
2.2.4. Gaussian Logistic Regression Model
2.2.5. Nonlinear Regression and Correlation Test
3. Results
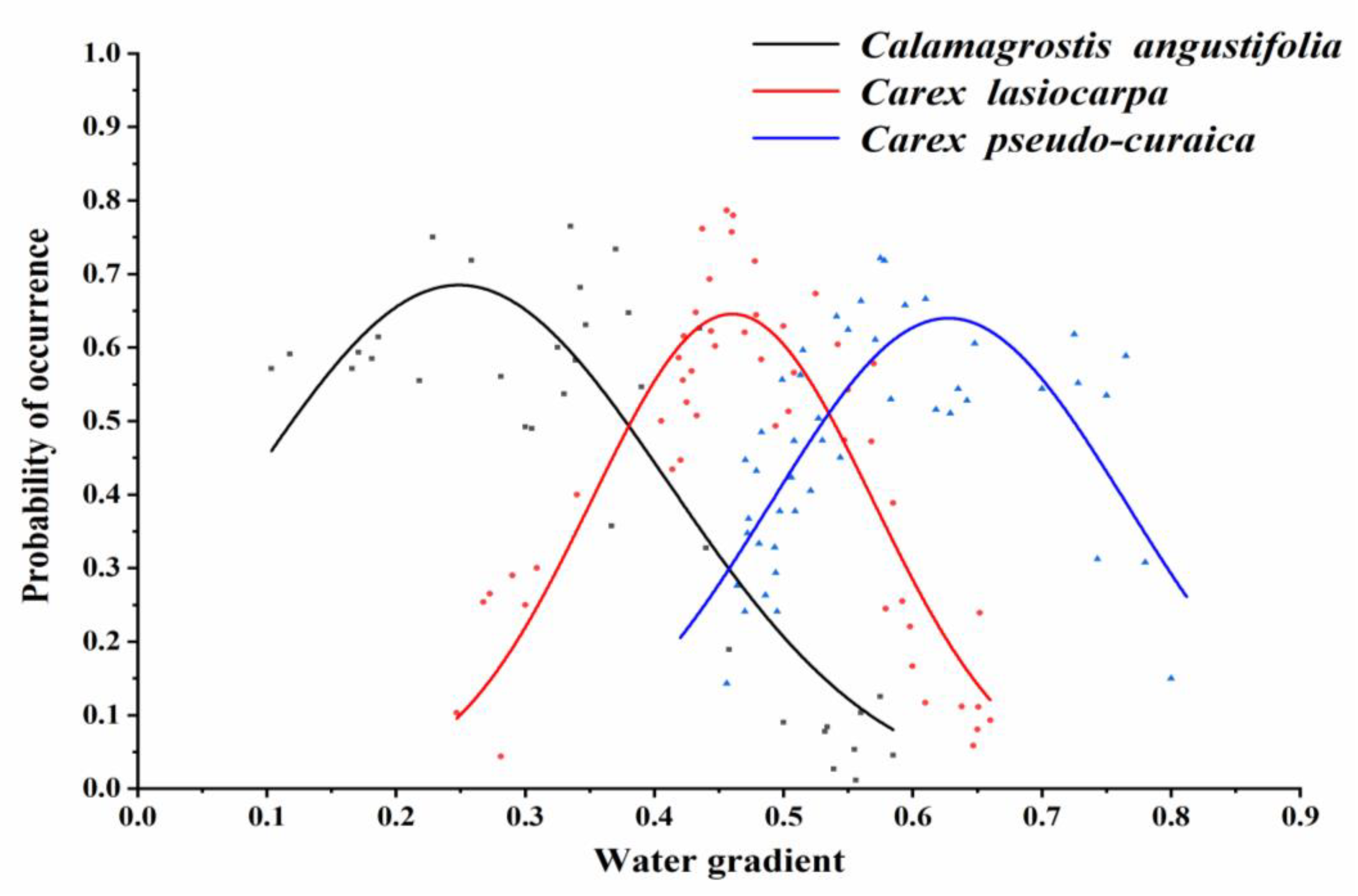
3.1. Species Response to the Hydrological Gradient
3.1.1. Response of Calamagrostis angustifolia to the Hydrological Gradient
3.1.2. Response of Carex lasiocarpa to the Hydrological Gradient
3.1.3. Response of Carex pseudocuraica to the Hydrological Gradient
3.2. Comparison of Probabilities of Occurrence Responses to the Hydrological Gradient
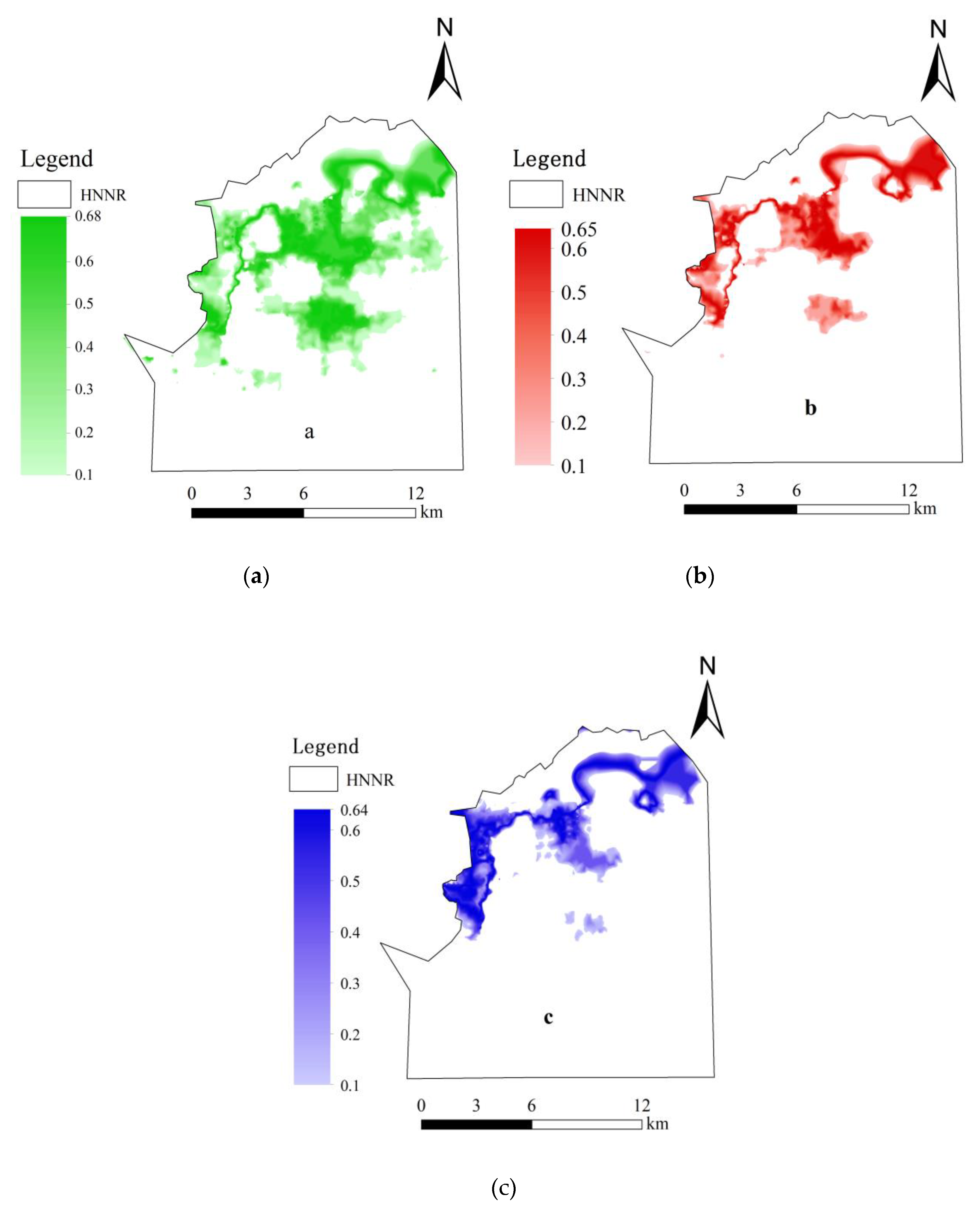
3.3. Simulation of the Spatial Distribution Probability
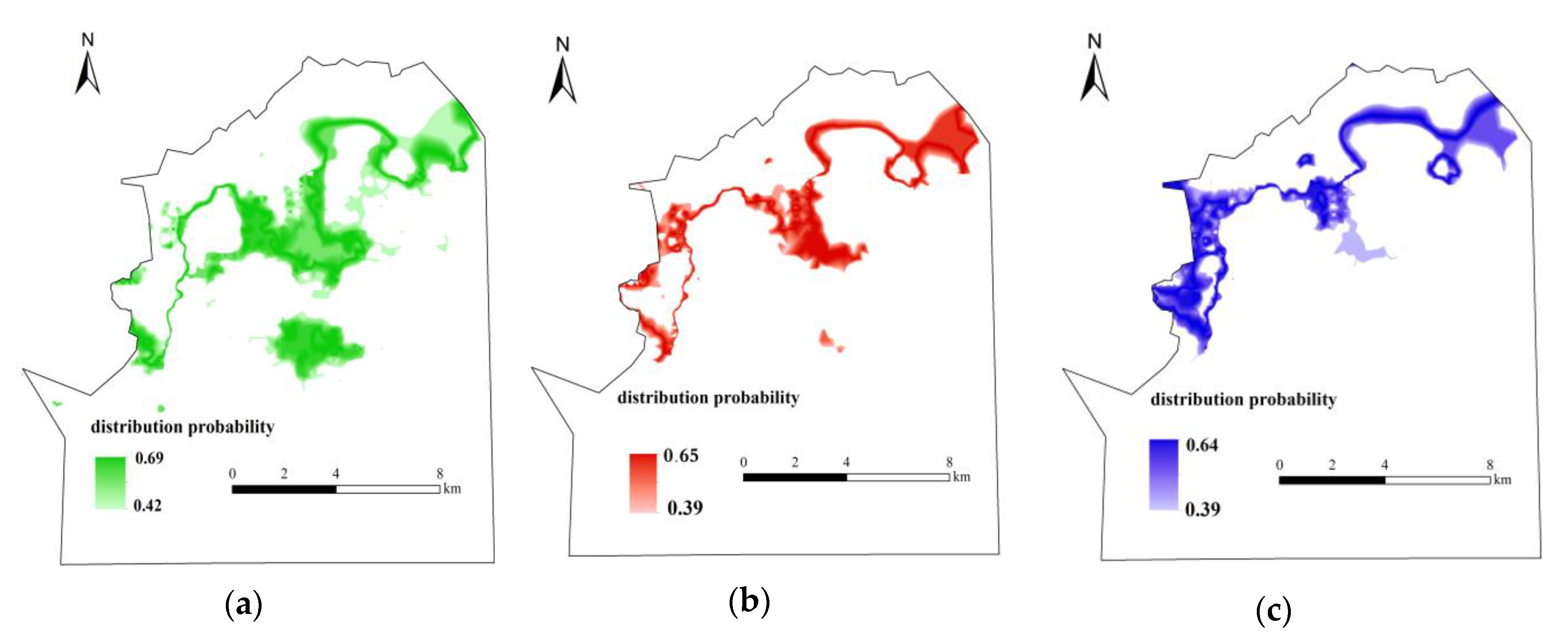
3.4. The Distribution of the Most Suitable Habitat
3.5. Validation Results
4. Discussion
4.1. Modeling the Spatial Distribution of Wetland Vegetation Species’ Response to the Hydrological Gradient
4.2. Uncertaintyof the Modeling the Spatial Distribution of Wetland Vegetation Species’ Response to the Hydrological Gradient
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foti, R.; Del Jesus, M.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydroperiod regime controls the organization of plant species in wetlands. Proc. Natl. Acad. Sci. USA 2012, 109, 19596–19600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinser, P.; Fox, S.; Keenan, L.; Ceric, A.; Baird, F. Hydrology and the Distribution of Floodplain Plant Communities of the Upper Set. Johns River, Florida—Using Transects to Track the Movement of Plant Communities Along a Changing Hydrological Gradient; St. Johns River Water Management District: Parateca, FL, USA, 2012. [Google Scholar]
- Todd, M.J.; Muneepeerakul, R.; Pumo, D.; Azaele, S.; Miralles-Wilhelm, F.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydrological drivers of wetland vegetation community distribution within Everglades National Park, Florida. Adv. Water Resour. 2010, 33, 1279–1289. [Google Scholar] [CrossRef]
- Wilcox, D.A.; Xie, Y.C. Predicting wetland plant community responses to proposed water-level-regulation plans for lake ontario: GIS-based modeling. J. Great Lakes Res. 2007, 33, 751–773. [Google Scholar] [CrossRef]
- Geest, G.J.V.; Coops, H.; Roijackers, R.M.M.; Buijse, A.D.; Scheffer, M. Succession of aquatic vegetation driven by reduced water-level fluctuations in floodplain lakes. J. Appl. Ecol. 2005, 42, 251–260. [Google Scholar] [CrossRef]
- Havens, K.E. Submerged aquatic vegetation correlations with depth and light attenuating materials in a shallow subtropical lake. Hydrobiologia 2003, 493, 173–186. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 2000, 147, 237–250. [Google Scholar] [CrossRef]
- Wang, H.Y.; Chen, J.K.; Zhou, J. Influence of water level gradiention plant growth, reproduction and biomass allocation of wetland plant species. Chin. J. Plant Ecol. 1999, 3, 269–274. (In Chinese) [Google Scholar]
- Magee, T.K.; Kentula, M.E. Response of wetland plant species to hydrologic conditions. Wetl Ecol. Manag. 2005, 13, 163–181. [Google Scholar] [CrossRef]
- Wang, Q.L.; Chen, J.R.; Liu, H.; Yi, L.Y.; Li, W.; Liu, F. The growth responses of two emergent plants to the water depth. Acta Hydro. Sin. 2012, 33, 583–587. (In Chinese) [Google Scholar]
- Lou, Y.J.; Wang, G.P.; Lu, X.G.; Jiang, M.; Zhao, K.Y. Zonation of plant cover and environmental factors in wetlands of the Sanjiang Plain, northeast China. Nord. J. Bot. 2013, 31, 748–756. [Google Scholar] [CrossRef]
- Engels, J.G.; Jensen, K. Patterns of wetland plant diversity along estuarine stress gradients of the Elbe (Germany) and Connecticut (USA) Rivers. Plant Ecol.Diver. 2009, 2, 301–311. [Google Scholar] [CrossRef]
- Zhou, D.; Gong, H.; Luan, Z.; Hu, J.; Wu, F. Spatial pattern of water controlled wetland communities on the Sanjiang Floodplain, Northeast China. Community Ecol. 2006, 7, 223–234. [Google Scholar] [CrossRef]
- Wilson, S.D.; Keddy, P.A. Plant Zonation on a Shoreline Gradient: Physiological Response Curves of Component Species. J. Ecol. 1985, 73, 851–860. [Google Scholar] [CrossRef]
- Shi, F.X.; Song, C.C.; Zhang, X.H.; Mao, R.; Guo, Y.D.; Gao, F.Y. Plant zonation patterns reflected by the differences in plant growth, biomass partitioning and root traits along a water level gradient among four common vascular plants in freshwater marshes of the Sanjiang Plain, Northeast China. Ecol. Eng. 2015, 81, 158–164. [Google Scholar] [CrossRef]
- Correa-Araneda, F.J.; Urrutia, J.; Soto-Mora, Y.; Figueroa, R.; Hauenstein, E. Effects of the hydroperiod on the vegetative and community structure of freshwater forested wetlands, Chile. J.Freshw. Ecol. 2012, 27, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.H.; Ren, B.; Li, F. Increased nutrient supply facilitates acclimation to high-water level in the marsh plant Deyeuxia angustifolia: The response of root morphology. Aquat. Bot. 2009, 91, 1–5. [Google Scholar] [CrossRef]
- Crain, C.M. Interactions between marsh plant species vary in direction and strength depending on environmental and consumer context. J. Ecol. 2008, 96, 166–173. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Van der Valk, A.G.; Moloney, K.A. The role of water depth and soil temperature in determining initial composition of prairie wetland coenoclines. Plant. Ecol. 1998, 138, 203–216. [Google Scholar] [CrossRef]
- Dwire, K.A.; Kauffman, J.B.; Baham, B.J.E. Plant biomass and species composition along an environmental gradient in montane riparian meadows. Oecologia 2004, 139, 309–317. [Google Scholar] [CrossRef]
- Violle, C.; Bonis, A.; Plantegenest, M.; Cudennec, C.; Damgaard, C.; Marion, B.; Le Coeur, D.; Bouzille, J.B. Plant functional traits capture species richness variations along a flooding gradient. Oikos 2011, 120, 389–398. [Google Scholar] [CrossRef]
- Coudun, C.; Gegout, J.C. The derivation of species response curves with gaussian logistic regression is sensitive to sampling intensity and curve characteristics. Ecol. Model. 2006, 199, 164–175. [Google Scholar] [CrossRef]
- Lee, K.H.; Lunetta, R.S. Wetland detection methods. In Wetland and Environmental Application of GIS; Lyon, J.G., McCarthy, J., Eds.; Lewis: New York, NY, USA, 1995; pp. 249–284. [Google Scholar]
- Trepel, M.; Dall'O', M.; Cin, L.D.; Wit, M.D.; Opitz, S.; Palmeri, L.; Persson, J.; Pieterse, N.; Timmermann, T.; Bendoricchio, G.; et al. Models for wetland planning, design and management. EcoSys Bd. 2000, 8, 93–137. [Google Scholar]
- Hebb, A.J.; Mortsch, L.D.; Deadman, P.J.; Cabrera, A.R. Modeling wetland vegetation community response to water-level change at Long Point, Ontario. J. Great Lakes Res. 2013, 39, 191–200. [Google Scholar] [CrossRef]
- Jiao, C.C.; Zhou, D.M. Modeling the spatial distribution of Carex pseudocuraica in a freshwater marsh, Northeast China. Wetlands 2014, 34, 267–276. [Google Scholar] [CrossRef]
- Luan, Z.Q.; Wang, Z.X.; Yan, D.D.; Liu, G.H.; Xu, Y.Y. The ecological response of Carex lasiocarpa community in the Riparian Wetlands to the environmental gradient of water depth in Sanjiang Plain, Northeast China. Sci. World J. 2013, 2013, 1–7. [Google Scholar]
- Lou, Y.J.; Gao, C.Y.; Pan, Y.W.; Xue, Z.S.; Liu, Y.; Tang, Z.H.; Jiang, M.; Lu, X.G.; Rydin, H. Niche modelling of marsh plants based on occurrence and abundance data. Sci. TotalEnviron. 2018, 616, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.J.; Pan, Y.W.; Gao, C.Y.; Jiang, M.; Lu, X.G.; Xu, Y.J. Response of plant height, species richness and aboveground biomass to flooding gradient along vegetation zones in floodplain wetlands, Northeast China. PLoS ONE 2016, 11, e0153972. [Google Scholar]
- Adam, E.; Mutanga, O.; Rugege, D. Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: A review. Wetl. Ecol. Manag. 2010, 18, 281–296. [Google Scholar] [CrossRef]
- Zhao, K.Y. Mires in China; Science Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Luo, W.B.; Song, F.B.; Xie, Y.H. Trade-off between tolerance to drought and tolerance to flooding in three wetland plants. Wetlands 2008, 28, 866–873. [Google Scholar] [CrossRef]
- Zhang, X.H.; Mao, R.; Gong, C.; Yang, G.S.; Lu, Y.Z. Effects of hydrology and competition on plant growth in a freshwater marsh of northeast China. J. Freshwater. Ecol. 2014, 29, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Luan, Z.Q.; Deng, W.; Zhu, B.G. Estimation of eco-environmental water consumption for Honghe national nature reserve wetlands. Arid Land Resour. Environ. 2004, 18, 59–63. [Google Scholar]
- Liu, X.T. Natural Environmental Changes and Ecological Protection in the Sanjiang Plain; Science Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Zhou, D.M.; Gong, H.L. Hydro-Ecological Modeling of the Honghe National Natural Reserve; China Environmental Science Press: Beijing, China, 2007. (In Chinese) [Google Scholar]
- Fu, P.Y. Clavis Plantarum Chinese Boreali-Orientalis; Science Press: Beijing, China, 1995. (In Chinese) [Google Scholar]
- Zhou, D.M.; Gong, H.L.; Wang, Y.Y.; Khan, S.B.; Zhao, K.Y. Driving forces for the marsh wetland degradation in the Honghe National Nature Reserve in Sanjiang plain, Northeast China. Environ. Model. Assess. 2009, 14, 101–111. [Google Scholar] [CrossRef]
- Wan, L.H.; Zhang, Y.W.; Zhang, X.Y.; Qi, S.Q.; Na, X.D. Comparison of land use/land cover change and landscape patterns in Honghe National Nature Reserve and the surrounding Jiansanjiang Region, China. Ecol. Indic. 2015, 51, 205–214. [Google Scholar] [CrossRef]
- Urban, K.E. Oscillating vegetation dynamics in a wet heathland. J. Veg.Sci. 2009, 16, 111–120. [Google Scholar] [CrossRef]
- Zhang, J.T. Quantitative Ecology; Science Press: Beijing, China, 2010. (In Chinese) [Google Scholar]
- Cui, B.S.; Zhao, X.S.; Yang, Z.F.; Tang, N.; Tan, X.J. The response of reed community to the environment gradient of water depth in the Yellow River Delta. Acta. Ecol. Sin. 2006, 3, 194–202. [Google Scholar] [CrossRef]
- Bi, Z.L.; Xiong, X.; Lu, F.; He, Q.; Zhao, X.S. Studies on ecological amplitude of reed to the environmental gradient of waterdepth. Shandong For. Sci.Technol. 2007, 4, 1–3. (In Chinese) [Google Scholar]
- Lou, Y.J.; Zhao, K.Y.; Wang, G.P.; Jiang, M.; Lu, X.G.; Rydin, H. Long term changes in marsh vegetation in Sanjiang plain, Northeast China. J. Veg. Sci. 2015, 26, 643–650. [Google Scholar] [CrossRef]
- Austin, M.P. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wetland Vegetation Species | Coefficient | Performance Evaluation Criteria (p < 0.05) | ||||||
|---|---|---|---|---|---|---|---|---|
| c | u | t | F-Statistic | R | R2 | RMSE | SSE | |
| Calamagrostis angustifolia | 0.68 | 0.25 | 0.16 | 140.4 | 0.90 | 0.81 | 0.11 | 0.40 |
| Carex lasiocarpa | 0.65 | 0.46 | 0.11 | 241.9 | 0.91 | 0.83 | 0.09 | 0.41 |
| Carex pseudocuraica | 0.62 | 0.63 | 0.14 | 59.88 | 0.76 | 0.58 | 0.10 | 0.42 |
| Transect | Wetland Vegetation Species | Performance Evaluation Criteria (p < 0.05) | ||
|---|---|---|---|---|
| R2 | SE | SEM | ||
| Transect 1 | Calamagrostis angustifolia | 0.62 | 0.24 | 0.11 |
| Carex lasiocarpa | 0.68 | 0.27 | 0.17 | |
| Carex pseudocuraica | ||||
| Transect 2 | Calamagrostis angustifolia | 0.84 | 0.14 | 0.03 |
| Carex lasiocarpa | 0.93 | 0.10 | 0.02 | |
| Carex pseudocuraica | 0.97 | 0.09 | 0.02 | |
| Transect 3 | Calamagrostis angustifolia | |||
| Carex lasiocarpa | 0.71 | 0.19 | 0.07 | |
| Carex pseudocuraica | 0.79 | 0.22 | 0.09 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, D.; Luan, Z.; Xu, D.; Xue, Y.; Shi, D. Modeling the Spatial Distribution of Three Typical Dominant Wetland Vegetation Species’ Response to the Hydrological Gradient in a Ramsar Wetland, Honghe National Nature Reserve, Northeast China. Water 2020, 12, 2041. https://doi.org/10.3390/w12072041
Yan D, Luan Z, Xu D, Xue Y, Shi D. Modeling the Spatial Distribution of Three Typical Dominant Wetland Vegetation Species’ Response to the Hydrological Gradient in a Ramsar Wetland, Honghe National Nature Reserve, Northeast China. Water. 2020; 12(7):2041. https://doi.org/10.3390/w12072041
Chicago/Turabian StyleYan, Dandan, Zhaoqing Luan, Dandan Xu, Yuanyuan Xue, and Dan Shi. 2020. "Modeling the Spatial Distribution of Three Typical Dominant Wetland Vegetation Species’ Response to the Hydrological Gradient in a Ramsar Wetland, Honghe National Nature Reserve, Northeast China" Water 12, no. 7: 2041. https://doi.org/10.3390/w12072041