On the Occurrence of Metadiaptomus chevreuxi (Calanoida, Diaptomidae, Paradiaptominae) in the Iberian Peninsula, With Notes on the Ecology and Distribution of its European Populations


Abstract
:1. Introduction
2. Materials and Methods
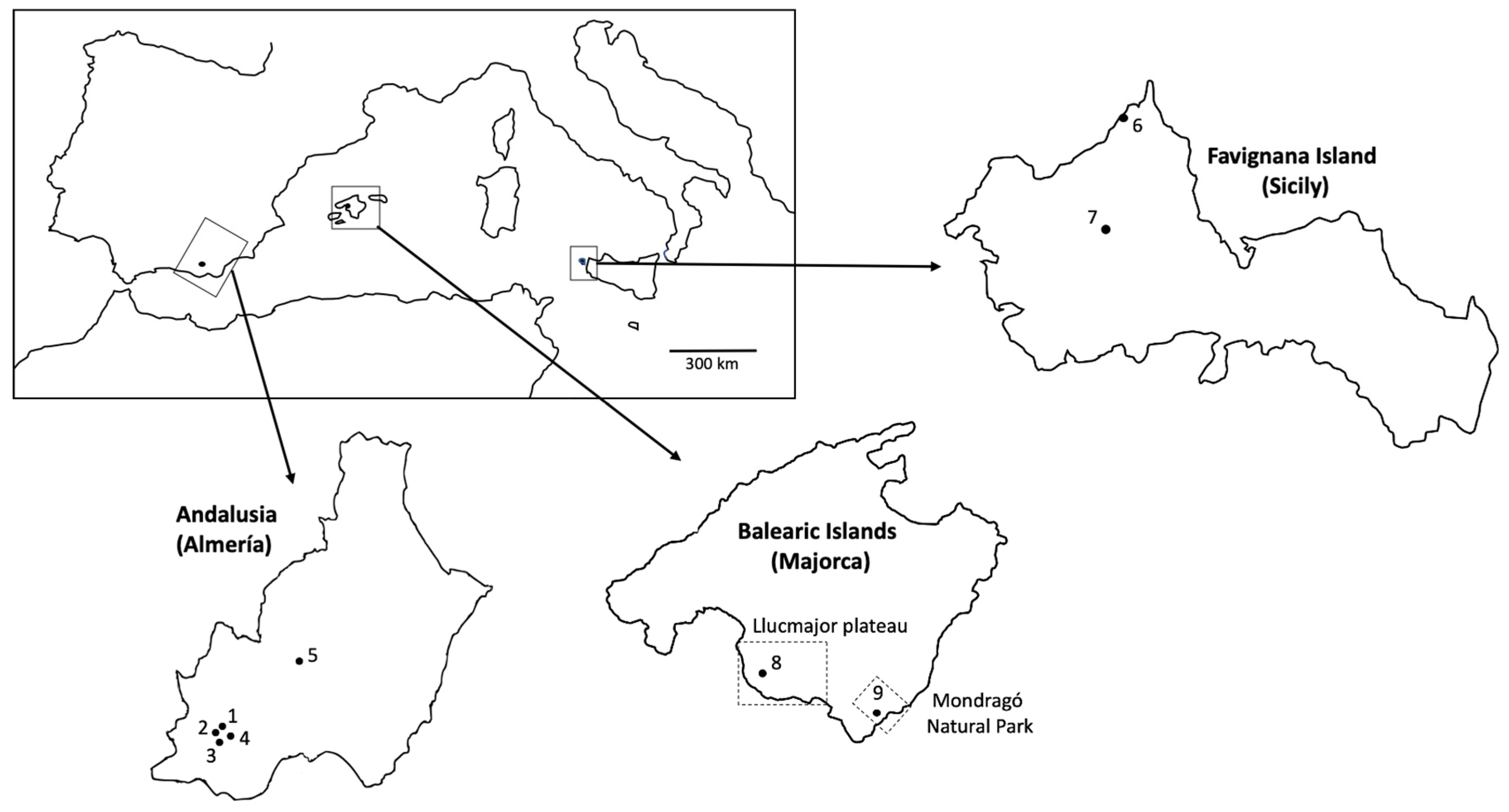
2.1. Study Areas
2.1.1. Andalusia
2.1.2. Sicily
2.1.3. Majorca
2.2. Sampling Methods and Crustacean Identification
2.2.1. Sampling
2.2.2. Crustacean Identification
3. Results
3.1. The Aquatic Environment
3.2. Co-Occurring Flora and Vertebrate Fauna
3.3. Crustacean Assemblages
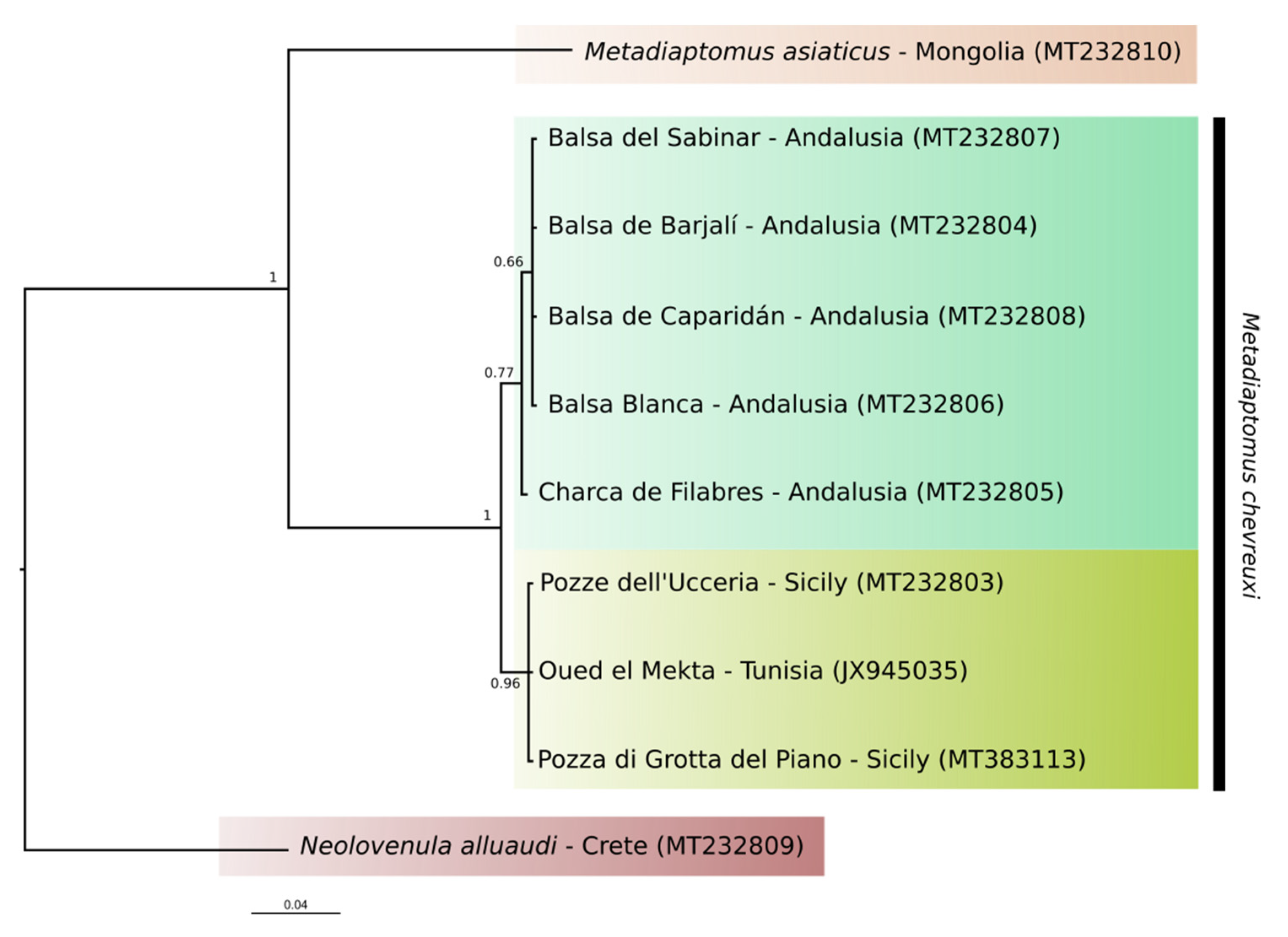
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miracle, M.R. Biogeography of the freshwater zooplanktonic communities of Spain. J. Biogeogr. 1982, 9, 455–467. [Google Scholar] [CrossRef]
- Baltanás, A.; Montes, C.; Martino, P. Distribution patterns of ostracods in Iberian saline lakes. Ecological factors. Hydrobiologia 1990, 197, 207–220. [Google Scholar] [CrossRef]
- Alonso, M. Las lagunas de la España peninsular. Limnetica 1998, 15, 1–176. [Google Scholar]
- Schmit, O.; Adolfsson, S.; Vandekerkhove, J.; Rueda, J.; Bode, S.; Rossetti, G.; Michalakis, Y.; Jokela, J.; Martens, K.; Mesquita-Joanes, F. The distribution of sexual reproduction of the geographic parthenogen Eucypris virens (Crustacea: Ostracoda) matches environmental gradients in a temporary lake. Can. J. Zoöl. 2013, 91, 660–671. [Google Scholar] [CrossRef]
- Sala, J.; Gascón, S.; Cunillera-Montcusí, D.; Alonso, M.; Amat, F.; Da Fonseca, L.C.; Cristo, M.; Florencio, M.; García, C.M.; Machado, M.; et al. Defining the importance of landscape metrics for large branchiopod biodiversity and conservation: The case of the Iberian Peninsula and Balearic Islands. Hydrobiologia 2017, 801, 81–98. [Google Scholar] [CrossRef]
- Mesquita-Joanes, F.; Aguilar-Alberola, J.A.; Palero, F.; Rueda, J. A new species of Cypris (Crustacea: Ostracoda) from the Iberian Peninsula and the Balearic Islands, with comments on the first ostracod named using the Linnean system. Zootaxa 2020, 4759, 113–131. [Google Scholar] [CrossRef]
- Dumont, H.J. Zooplankton and the Science of Biogeography: The Example of Africa. In Evolution and Ecology of Zooplankton Communities; Kerfoot, W.C., Ed.; University Press of New England: Lebanon, NH, USA, 1980; pp. 685–696. [Google Scholar]
- Leibold, M.A.; Economo, E.P.; Neto, P.P. Metacommunity phylogenetics: Separating the roles of environmental filters and historical biogeography. Ecol. Lett. 2010, 13, 1290–1299. [Google Scholar] [CrossRef]
- Marrone, F.; Brutto, S.L.; Hundsdoerfer, A.K.; Arculeo, M. Overlooked cryptic endemism in copepods: Systematics and natural history of the calanoid subgenus Occidodiaptomus Borutzky 1991 (Copepoda, Calanoida, Diaptomidae). Mol. Phylogenetics Evol. 2013, 66, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Margalef, R. Los Crustáceos de las Aguas Continentales Ibéricas; Instituto Forestal de Investigaciones y Experiencias, Ministerio de Agricultura: Madrid, Spain, 1953; pp. 1–243. [Google Scholar]
- Ortiz, E. Diaptómidos de las aguas continentales españolas (Crustacea, Copepoda). Bol. Real. Soc. Esp. Hist. Nat. Sec. Biol. 1957, 55, 391–398. [Google Scholar]
- Alonso, M. The genus Mixodiaptomus Kiefer, 1932 (Copepoda, Diaptomidae) in Spain. Hydrobiologia 1984, 118, 135–146. [Google Scholar] [CrossRef]
- Caramujo, M.J.; Boavida, M.-J. Biological diversity of copepods and cladocerans in Mediterranean temporary ponds under periods of contrasting rainfall. J. Limnol. 2010, 69, 64. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.D.; De Vicente, I.; Ortega, F.; Jiménez-Melero, R.; Parra, G.; Guerrero, F.; Ruiz, J. A comprehensive evaluation of the crustacean assemblages in southern Iberian Mediterranean wetlands. J. Limnol. 2015, 73, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Fidalgo, M.L.; Monteiro, M.T. A Note on the Presence of Dussartius Baeticus (Dussart, 1967) in Portugal (Copepoda, Calanoida). Crustaceana 2003, 76, 1399–1407. [Google Scholar] [CrossRef]
- Vecchioni, L.; Sala, J.; Arculeo, M.; Marrone, F. On the occurrence of Eudiaptomus gracilis (G.O. Sars, 1863) (Copepoda: Diaptomidae) in Madeira (Portugal). Limnetica 2020, 39, 571–578. [Google Scholar] [CrossRef]
- Sahuquillo, M.; Miracle, M.R. The role of historic and climatic factors in the distribution of crustacean communities in Iberian Mediterranean ponds. Freshw. Biol. 2013, 58, 1251–1266. [Google Scholar] [CrossRef]
- Taglialatela, S.; Ruiz, J.; Prieto, L.; Navarro, G. Seasonal forcing of image-analysed mesozooplankton community composition along the salinity gradient of the Guadalquivir estuary. Estuar. Coast. Shelf Sci. 2014, 149, 244–254. [Google Scholar] [CrossRef]
- Dussart, B.; Defaye, D. World Directory of Crustacea Copepoda of Inland Waters. I Calaniformes; Backuhys Publishers: Leiden, The Netherlands, 2002; pp. 1–276. [Google Scholar]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe. Cladocera & Copepoda (Calanoida, Cyclopoida) Key to Species Identification, with Notes on Ecology, Distribution, Methods and Introduction to Data Analysis; Springer: Basel, Switzerland, 2016; pp. 1–918. [Google Scholar]
- Marrone, F.; Alfonso, G.; Naselli-Flores, L.; Stoch, F. Diversity patterns and biogeography of Diaptomidae (Copepoda, Calanoida) in the Western Palearctic. Hydrobiologia 2017, 800, 45–60. [Google Scholar] [CrossRef]
- Sanz de Galdeano, C.; López-Garrido, A. Structure of the Sierra de Lujar (Alpujarride Complex, Betic Cordillera). Estud. Geológicos 2014, 70, 005. [Google Scholar] [CrossRef]
- Villalobos, M. Geología y Geomorfología. In Sierra de Gádor. Patrimonio Natural e Infraestructura Verde de Almería; Oyonarte, C., Giménez, E., Villalobos, M., Guirado, J., Eds.; Fundación Patrimonio Natural, Biodiversidad y Cambio Global: Almería, Spain, 2016; pp. 41–56. [Google Scholar]
- Corbel, J. Les Karst du Nord-Ouest l’Europe; Institut des Études Rhodaniennes de l’Université de Lyon: Lyon, France, 1957; pp. 1–544. [Google Scholar]
- Sanz De Galdeano, C.; López-Garrido, A. The nevado-filábride complex in the western part of Sierra de los Filabres (Betic Internal Zone), structure and lithologic succession. Boletín Geológico Min. 2016, 127, 823–836. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Zotano, J.; Alcántara-Manzanares, J.; Olmedo-Cobo, J.A.; Martínez-Ibarra, E. La sistematización del clima mediterráneo: Identificación, clasificación y caracterización climática de Andalucía (España). Rev. Geogr. Norte Gd. 2015, 61, 161–180. [Google Scholar] [CrossRef] [Green Version]
- S.A.I.H. HIDROSUR. Available online: http://www.redhidrosurmedioambiente.es/saih/resumen/precipitacion/historica (accessed on 6 April 2020).
- Marrone, F.; Naselli-Flores, L. First record of a representative of the subfamily Paradiaptominae (Copepoda Calanoida Diaptomidae) in Italy: Metadiaptomus chevreuxi (Guerne & Richard, 1894). J. Limnol. 2005, 64, 89–92. [Google Scholar] [CrossRef]
- Pepe, F.; Corradino, M.; Parrino, N.; Besio, G.; Presti, V.L.; Renda, P.; Calcagnile, L.; Quarta, G.; Sulli, A.; Antonioli, F. Boulder coastal deposits at Favignana Island rocky coast (Sicily, Italy): Litho-structural and hydrodynamic control. Geomorphology 2018, 303, 191–209. [Google Scholar] [CrossRef]
- Zampino, D.; Duro, A.; Piccione, V.; Scalia, C. Fitoclima della Sicilia. Termoudogrammi Secondo Walter e Lieth. In Atti V Workshop Progetto Strategico Clima, Ambiente e Territorio nel Mezzogiorno; Guerrini, E., Ed.; C.N.R.: Roma, Italy, 1997; Volume II, pp. 7–54. [Google Scholar]
- Jaume, D. Metadiaptomus chevreuxi (Copepoda: Diaptomidae) and Leptestheria mayeti (Branchiopoda: Conchostraca: Leptestheriidae), two African freshwater crustaceans recorded in Majorca. Limnetica 1989, 5, 101–109. [Google Scholar]
- Jaume, D. Calanoides (Crustacea: Copepoda) de las aigües continentals baleàriques. Boll. Soc. Hist. Nat. Balears 1989–1990, 33, 207–219. [Google Scholar]
- Pinya, S.; Gil, L. Primary Results on Biodiversity Analysis of the Temporary Ponds in the Natural Park of Mondragó, Mallorca. In International Conference on Mediterranean Temporary Ponds. Proceedings & Abstracts; Fraga, I., Argiumbau, P., Eds.; Consell Insular de Menorca: Menorca, Spain, 2009; pp. 291–295. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 23th ed.; American Public Health Association: Washington, DC, USA, 2017; pp. 1–1540. [Google Scholar]
- Cirujano, S.; Cambra, J.; Sánchez-Castillo, P.M.; Meco, A.; Flor-Arnau, N. Flora Ibérica. Algas Continentales. Carófitos (Characeae); Real Jardín Botánico: Madrid, Spain, 2008; pp. 1–132. [Google Scholar]
- Cirujano, S.; Meco-Molina, A.; García-Murillo, P. Flora Acuática Española. Hidrófitos Vasculares; Real Jardín Botánico-CSIC: Madrid, Spain, 2014; pp. 1–320. [Google Scholar]
- Parrondo-Celdrán, P. Guía de los Anfibios del Sureste Ibérico; Asociación Columbares: Murcia, Spain, 2013; pp. 1–39. [Google Scholar]
- Ambrogio, A.; Mezzadri, S. Girini d’Italia—Tadpoles of Italy; Societas Herpetologica Italica: Italy, 2014; pp. 1–102. [Google Scholar]
- Alonso, M. Crustacea, Branchiopoda. In Fauna Iberica; Ramos Sánchez, M.A., Tercedor, J.A., Bellés I Ros, X., Gosálbez i Noguera, J., Guerra Sierra, A., Macpherson Mayol, E., Piera, F.M., Serrano Marino, J., Templado González, J., Eds.; Museo Nacional de Ciencias Naturales CSIC: Madrid, Spain, 1996; Volume 7, pp. 1–486. [Google Scholar]
- Kiefer, F. Das Zooplankton der Binnengewässer. Freilebende Copepoda. Die Binnengewässer; E. Schweizerbart’sche Verlagbuchhandlung: Stuttgart, Germany, 1978; Volume 26, Part 2; pp. 1–343. [Google Scholar]
- Rayner, N.A. Copepoda: Calanoida (Diaptomidae: Paradiaptominae). Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Backhuys Publishers: Leiden, The Netherlands, 1999; pp. 1–122. [Google Scholar]
- Meisch, C. Freshwater Ostracoda of Western and Central Europe; Spektrum Akademischer Verlag GmbH: Heidelberg/Berlin, Germany, 2000; pp. 1–522. [Google Scholar]
- Marrone, F.; Alfonso, G.; Stoch, F.; Pieri, V.; Alonso, M.; Dretakis, M.; Naselli-Flores, L. An account on the non-malacostracan crustacean fauna from the inland waters of Crete, Greece, with the synonymization of Arctodiaptomus piliger Brehm, 1955 with Arctodiaptomus alpinus (Imhof, 1885) (Copepoda: Calanoida). Limnetica 2019, 38, 167–187. [Google Scholar] [CrossRef]
- Marrone, F.; Alonso, M.; Pieri, V.; Augugliaro, C.; Stoch, F. The crustacean fauna of Bayan Onjuul area (Tôv Province, Mongolia) (Crustacea: Branchiopoda, Copepoda, Ostracoda). North West. J. Zool. 2015, 11, 288–295. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Boil. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.E.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuyfzand, P.J. A New Hydrochemical Classification of Watertypes. In Regional Characterization of Water Quality. Proceedings of the Baltimore Symposium; Stephen, R., Ed.; IAHS Publ. 182: Washington, DC, USA, 1989; pp. 89–98. [Google Scholar]
- Rita, J.; Bibiloni, G. Zonación de la vegetación hidrófila de balsas periódicas en las zonas semiáridas de Baleares. Orsis 1991, 6, 61–74. [Google Scholar]
- Alemany, G.; Bauzá, A.; Cox, J.R.; Garcies, D.; Sampaio, H.S. Uso Público en el Parque Natural de Mondragó (Mallorca): Régimen Jurídico, Análisis de la Oferta y la Demanda y Propuestas de Actuación. In Turismo y Transformaciones Urbanas en el Siglo XXI; Pumares, P., Asensio, M.A., Fernández, F., Eds.; Universidad de Almería: Almería, Spain, 2002; pp. 149–162. [Google Scholar]
- Martínez-Solano, I. El sapo balear (Bufotes balearicus) en las Islas Baleares. Bol. Asoc. Herp. Esp. 2015, 26, 21–23. [Google Scholar]
- Altinsaçli, S.; Griffiths, H.I. The freshwater ostracods Hungarocypris and Leucocythere from Turkey. Crustaceana 2001, 74, 681–688. [Google Scholar] [CrossRef]
- Martínez-García, B.; Huerta, P.; Armenteros, I.; Rodríguez-Lázaro, J. Distribución y ecología de los ostrácodos recientes de las Lagunas de Villafáfila (Zamora, España). Geogaceta 2019, 66, 75–78. [Google Scholar]
- Alfonso, G.; Belmonte, G. Neolovenula alluaudi (Guerne and Richard, 1890) (Calanoida: Diaptomidae: Paradiaptominae): First record in Italy and review of geographical distribution. J. Limnol. 2013, 72, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Gurney, R. Freshwater Crustacea collected by Dr. P.A. Buxton in Mesopotamia and Persia. J. Bombay Nat. Hist. Soc. 1921, 27, 85–93. [Google Scholar]
- Brehm, V. Contribution à l’étude de la faune d’Afghanistan. Nr. 17. Cladoceren und calanoide Kopepoden aus Afghanistan, nebst Beschreibung eines neuen Metadiaptomus aus Iran und Bemerkungen über einige andere Calanoiden aus Iran. SB Osterr. Akad. Wiss. Abt. I Math Nat Kl. 1959, 168, 917–935. [Google Scholar]
- Dumont, H.J. Limnologie van Sahara en Sahel. Ph.D. Thesis, University of Ghent, Gent, Belgium, 1979. [Google Scholar]
- Miracle, M.R.; Sahuquillo, M.; Vicente, E. Large branchiopods from freshwater temporary ponds of Eastern Spain. Verh. Internat. Ver. Limnol. 2008, 30, 501–505. [Google Scholar] [CrossRef]
- Alfonso, G. Diversity and distribution of large branchiopods (Branchiopoda: Anostraca, Notostraca, Spinicaudata) in Apulian ponds (SE Italy). Eur. Zool. J. 2017, 84, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, H. Recherches Sur le Faune des Eaux Continentales de l’Algérie et de la Tunisie; Thése Sciences, Imprimerie Minerva: Alger, Tunisia, 1928; pp. 1–419. [Google Scholar]
- Kiefer, F. Freilebende Ruderfuβkrebse (Crustacea, Copepoda) aus Binnengewässern Marokkos. Bull. Soc. Sc. Nat. Phys. Maroc 1954, 34, 317–336. [Google Scholar]
- Turki, S.; El Abed, A. Nouvelles informations sur les copépodes calanoïdes et cyclopoïdes des eaux continentals tunisiennes. Crustaceana 1999, 72, 157–169. [Google Scholar] [CrossRef]
- Leibowitz, S.G. Isolated wetlands and their functions: An ecological perspective. Wetlands 2003, 23, 517–531. [Google Scholar] [CrossRef]
- Sainz-Ollero, H.; van Staalduinen, M.A. Iberian Steppes. In Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World; Werger, M.J.A., van Staalduinen, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 273–288. [Google Scholar] [CrossRef]
- Sáez, L.; Fraga, P.; López-Alvarado, J. The Flora of the Balearic Islands. In Islands and Plants: Preservation and Understanding of Flora on Mediterranean Islands. 2nd Botanical Conference in Menorca; Cardona, E., Estaún, I., Comas, M., Fraga, P., Blanché, C., Eds.; Institut Menorquí d’Estudis: Maó, Spain, 2013; pp. 91–103. [Google Scholar] [CrossRef]
- Guarino, R.; Pasta, S. Botanical Excursions in Central and Western Sicily. Field Guide for the 60th IAVS Symposium Palermo, 20–24 June 2017; Palermo University Press: Palermo, Italy, 2017; pp. 1–604. [Google Scholar]
- Robach, F.; Thiébaut, G.; Trémolières, M.; Müller, S. A reference system for continental running waters: Plant communities as bioindicators of increasing eutrophication in alkaline and acidic waters in north-east France. Hydrobiologia 1996, 340, 67–76. [Google Scholar] [CrossRef]
- Cirujano, S.; Medina, L. Plantas Acuáticas de las Lagunas y Humedales de Castilla-La Mancha; Junta de Comunidades de Castilla-La Mancha-CSIC: Madrid, Spain, 2002; pp. 1–336. [Google Scholar]
- Álvarez, J.; Salvador, A.; Martín, J.; López, P. Desarrollo larvario del sapo corredor, Bufo calamita (Laurenti, 1768) (Anura, Bufonidae) en charcas temporales del noreste español. Bol. R. Soc. Esp. Hist. Nat. Sec. Biol. 1992, 88, 129–137. [Google Scholar]
- De Pablo, F. Programa de Seguimiento de Anfibios en la Reserva de Biosfera de Menorca. Año 2016. Informe Técnico 01/2017; Departament de Medi Ambient i Reserva de Biosfera, Consell Insular de Menorca: Maón, Spain, 2016. [Google Scholar]
- Roca, J.R.; Mezquita, F.; Rueda, J.; Camacho, A.; Miracle, M.R. Endorheic versus karstic lakes: Patterns of ostracod distributions and lake typology in a Mediterranean landscape (Castilla-La Mancha, Spain). Mar. Freshw. Res. 2000, 51, 311. [Google Scholar] [CrossRef]
- Brehm, V. Reflexiones sobre relaciones zoogeográficas de la fauna de agua dulce de la Península Ibérica. Publ. Inst. Biol. Apl. 1947, 4, 53–74. [Google Scholar]
- Margalef, R. Estudios sobre la vida en las aguas continentales de la región endorreica manchega. Publ. Inst. Biol. Apl. 1947, 4, 5–51. [Google Scholar]
- Marrone, F.; Castelli, G.; Naselli-Flores, L. Sicilian Temporary Ponds: An Overview on the Composition and Affinities of their Crustacean Biota. In International Conference on Mediterranean Temporary Ponds. Proceedings & Abstracts; Fraga, I., Argiumbau, P., Eds.; Consell Insular de Menorca: Menorca, Spain, 2009; pp. 189–202. [Google Scholar]
- Champeau, A.; Thiéry, A. Les Crustacés Entomostracés des eaux stagnantes de Corse. Importance particulière des espèces monovoltines méditerranéennes de Copépodes Calanoïdes et d’Anostracés dans le Sud-Est de l’île. Bull. Soc. Zool. Fr. 1990, 115, 55–75. [Google Scholar]
- Alonso, M. Ejemplos de vicarianza en comunidades de crustáceos de laguna esteparias. Limnetica 1987, 3, 81–89. [Google Scholar]
- Downing, J.A. Emerging global role of small lakes and ponds: Little things mean a lot. Limnetica 2010, 29, 9–24. [Google Scholar] [CrossRef]
- Paton, P.W.C. A review of vertebrate community composition in seasonal forest pools of the northeastern United States. Wetl. Ecol. Manag. 2005, 13, 235–246. [Google Scholar] [CrossRef]
- Colburn, E.A.; Weeks, S.C.; Reed, S.K. Diversity and Ecology of Vernal Pool Invertebrates. In Science and Conservation of Vernal Pools in Northeastern North America; Calhoun, A.J.K., de Maynadier, P.G., Eds.; CRC: New York, NY, USA, 2008; pp. 105–126. [Google Scholar] [CrossRef]
- Mitchell, J.C.; Paton, P.W.C.; Raithel, C.J. The Importance of Vernal Pools to Reptiles, Birds, and Mammals. In Science and Conservation of Vernal Pools in Northeastern North America; Calhoun, A.J.K., deMaynadier, P.G., Eds.; CRC: New York, NY, USA, 2008; pp. 169–190. [Google Scholar] [CrossRef]
- Hassall, C.; Hollinshead, J.; Hull, A. Environmental correlates of plant and invertebrate species richness in ponds. Biodivers. Conserv. 2011, 20, 3189–3222. [Google Scholar] [CrossRef]
- Gounand, I.; Harvey, E.; Little, C.J.; Altermatt, F. Meta-Ecosystems 2.0: Rooting the Theory into the Field. Trends Ecol. Evol. 2018, 33, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M.; Mouquet, N.; Holt, R.D. Meta-ecosystems: A theoretical framework for a spatial ecosystem ecology. Ecol. Lett. 2003, 6, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Ortega, F.; Paracuellos, M.; Guerrero, F. Corología de macrófitos acuáticos en Andalucía oriental. Lazaroa 2004, 25, 179–185. [Google Scholar]
- Cirujano, S.; Guerrero, N.; García-Murillo, P. The genus Tolypella (A. Braun) A. Braun in the Iberian Peninsula. Acta Bot. Gall. 2013, 160, 121–129. [Google Scholar] [CrossRef]
- Blanco, S.; Olenici, A.; Ortega, F.; Jiménez-Gómez, F.; Guerrero, F. Taxonomía y morfología de Craticula gadorensis sp. nov. (Bacillariophyta, Stauroneidaceae). Bol. Soc. Arg. Bot. 2019, 54, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Blanco, S.; Olenici, A.; Jiménez-Gómez, F.; Ortega, F.; Guerrero, F. Una nueva especie del género Hantzschia (Bacillariaceae) en Almería, España. Caldasia 2019, 41, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Giménez, E.; Delgado, I.C.; Gómez Mercado, F. Comunidades vegetales de las balsas de la Sierra de Gádor (Almería). Lazaroa 2006, 27, 79–88. [Google Scholar]
- López-Medina, M.J. Las Balsas de Sierra de Gádor: Humedales en el Extremo Oriental de la Bética. In Lacus Autem Idem, et Stagnus, ubi Inmensa Aqua Convenit. Estudios Históricos Sobre Humedales en la Bética (II); Lagóstena, L.G., Ed.; Seminario Agustín de Horozco de Estudios Económicos de Historia Antigua y Medieval: Cádiz, Spain, 2016; pp. 15–43. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Blanca | Caparidán | Sabinar | Barjalí | Filabres | Pozze dell’Ucceria | Pozza di Grotta del Piano | |
|---|---|---|---|---|---|---|---|
| Coordinates (latitude) | 36°56′49.55″ N | 36°55′53.70″ N | 36°52′56.11″ N | 36°55′8.30″ N | 37° 8′21.10″ N | 37°56′58.71” N | 37°56′1.25′’ N |
| Coordinates (longitude) | 2°50′13.84″ W | 2°54′17.48″ W | 2°51′35.97″ W | 2°47′53.79″ W | 2°30′49.55″ W | 12°18′5.17′’ E | 12°17′59.41′’ E |
| Country/region | Spain/Andalusia | Spain/Andalusia | Spain/Andalusia | Spain/Andalusia | Spain/Andalusia | Italy/Sicily | Italy/Sicily |
| Municipality | Fondón | Laujar de Andarax | Berja-Dalías | Dalías-Padules-Almócita | Gérgal | Favignana | Favignana |
| Sampling date | 11/05/2018 | 11/05/2018 | 11/05/2018 | 11/05/2018 | 30/04/2018 | 8/04/2018 | 8/04/2018 |
| Included in a Nature 2000 site | SAC ES6110008 | SAC ES6110008 | SAC ES6110008 | SAC ES6110008 | - | SAC ITA010004 | SAC ITA010004 |
| Altitude (m a.s.l.) | 1551 | 1601 | 1830 | 1713 | 1050 | 3 | 30 |
| Maximum dimensions (m) | 13 × 14 | 67 × 30 | 85 × 70 | 74 × 50 | 17 × 14 | 95 × 16 | 8 × 3 |
| Hydroperiod | Temporary | Temporary | Temporary 1 | Temporary 1 | Temporary | Temporary | Temporary |
| Conductivity (μS cm−1) | 170 | 200 | 120 | 100 | 150 | 1510 | 316 |
| Turbidity (NTU) | 56 | 228 | 238 | 155 | 52 | 466 | 0.14 |
| pH | 8.90 | 8.50 | 8.95 | 9.43 | 9.59 | n.a. | n.a. |
| Alkalinity (mg L−1) | 74 | 94 | 55 | 49 | 59 | 180 | 60 |
| Nitrate (N-NO3− mg L−1) | 0.016 | <0.002 | <0.002 | <0.002 | 0.51 | 1.07 | 0.41 |
| Ammonia (N-NH4+ mg L−1) | <0.008 | <0.008 | <0.008 | <0.008 | 0.031 | 1.41 | n.a. |
| Phosphates (P-PO43− mg L−1) | <0.003 | <0.003 | <0.003 | 0.026 | <0.003 | 0.23 | 0.1 |
| Total Nitrogen (mg L−1) | 1.2 | 1.8 | 1.7 | 1.8 | 2.3 | 8.3 | 1.7 |
| Total Phosphorus (mg L−1) | 0.1 | 0.2 | 0.3 | 0.3 | <0.1 | 0.6 | 0.1 |
| Vegetation | I, II, III | I, II, III, IV | I, II, III, IV | I, II, III, IV | I *, II, III | - | V, VI |
| Vertebrate Fauna | VII | VII | VII | VII | VII | - | VIII |
| Taxa | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| COPEPODA | |||||||
| Calanoida | |||||||
| Metadiaptomus chevreuxi (Guerne & Richard, 1894) | X | X | X | X | X | X | X |
| Cyclopoida | |||||||
| Metacyclops minutus (Claus, 1863) | X | X | X | X | |||
| Cyclopoida n.i. | X 4 | ||||||
| BRANCHIOPODA | |||||||
| Anostraca | |||||||
| Branchipus schaefferi Fischer, 1834 | X | X | X | X | |||
| Notostraca | |||||||
| Triops cancriformis (Bosc, 1801) | X | ||||||
| Anomopoda | |||||||
| Daphnia (Ctenodaphnia) atkinsoni Baird, 1859 | X | X | X | X | X | X | X |
| Ceriodaphnia sp. 1 | X 1 | X 1 | |||||
| Ceriodaphnia sp. 2 | X 6 | ||||||
| Coronatella (Ephemeralona) orellanai (Alonso, 1996) | X | X | X | X | X | ||
| Coronatella (Ephemeralona) elegans (Kurz, 1875) | X | X | |||||
| Pleuroxus letourneuxi (Richard, 1888) | X | ||||||
| Macrothrix hirsuticornis Norman & Brady, 1867 | X | ||||||
| OSTRACODA | |||||||
| Podocopa | |||||||
| Eucypris virens (Jurine, 1820) | X | X | X 1 | X 2 | X 3 | X | X |
| Tonnacypris lutaria (Koch, 1838) | X | ||||||
| Heterocypris incongruens (Ramdohr, 1808) | X 5 | ||||||
| Heterocypris cf. barbara (Gauthier & Brehm, 1928) | X | X | |||||
| Plesiocypridopsis newtoni (Brady & Robertson, 1870) | X | ||||||
| Cypridopsis elongata (Kaufmann, 1900) | X | X | X | X | |||
| Potamocypris arcuata (Sars, 1903) | X | X | X | X | X | ||
| Cyprididae n.i. | X 4 | ||||||
| Leucocythere cf. mirabilis Kaufmann, 1892 | X |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marrone, F.; Ortega, F.; Mesquita-Joanes, F.; Guerrero, F. On the Occurrence of Metadiaptomus chevreuxi (Calanoida, Diaptomidae, Paradiaptominae) in the Iberian Peninsula, With Notes on the Ecology and Distribution of its European Populations. Water 2020, 12, 1989. https://doi.org/10.3390/w12071989
Marrone F, Ortega F, Mesquita-Joanes F, Guerrero F. On the Occurrence of Metadiaptomus chevreuxi (Calanoida, Diaptomidae, Paradiaptominae) in the Iberian Peninsula, With Notes on the Ecology and Distribution of its European Populations. Water. 2020; 12(7):1989. https://doi.org/10.3390/w12071989
Chicago/Turabian StyleMarrone, Federico, Fernando Ortega, Francesc Mesquita-Joanes, and Francisco Guerrero. 2020. "On the Occurrence of Metadiaptomus chevreuxi (Calanoida, Diaptomidae, Paradiaptominae) in the Iberian Peninsula, With Notes on the Ecology and Distribution of its European Populations" Water 12, no. 7: 1989. https://doi.org/10.3390/w12071989