Effects of Mixed Allelochemicals on the Growth of Microcystis aeruginosa, Microcystin Production, Extracellular Polymeric Substances, and Water Quality

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Microcystis Aeruginosa
2.2. Preparation of Allelochemicals and Their Combinations
2.3. Algal Inhibition Testing of Allelochemicals and Their Combinations
2.4. Measurements of Extracellular MC-LR Concentrations
2.5. Extraction and Characterization of EPS
2.6. Water Quality Assays
2.7. Statistical Analysis
3. Results
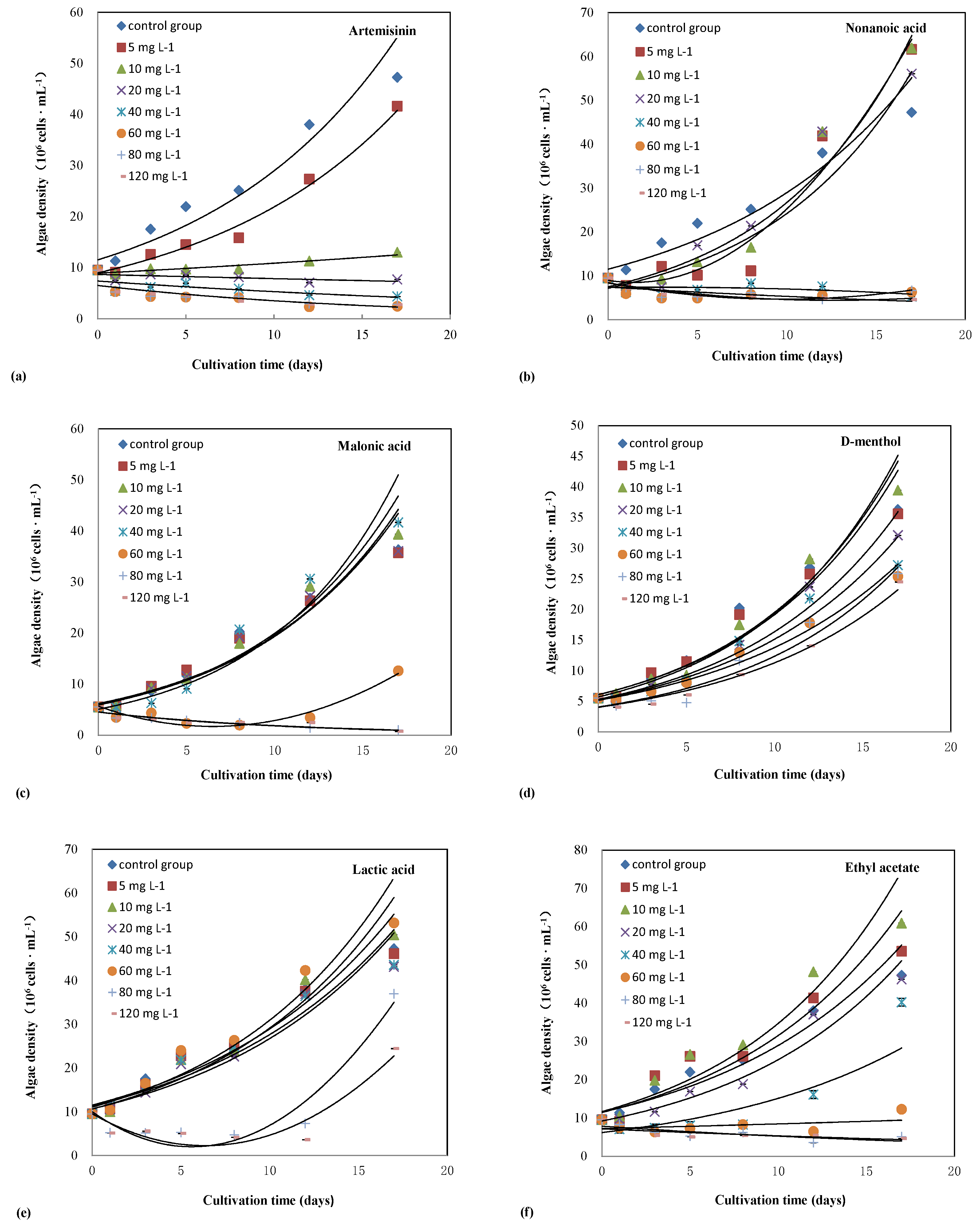
3.1. Inhibitory Effects of Six Different Chemicals on the Cell Density of Microcystis Aeruginosa
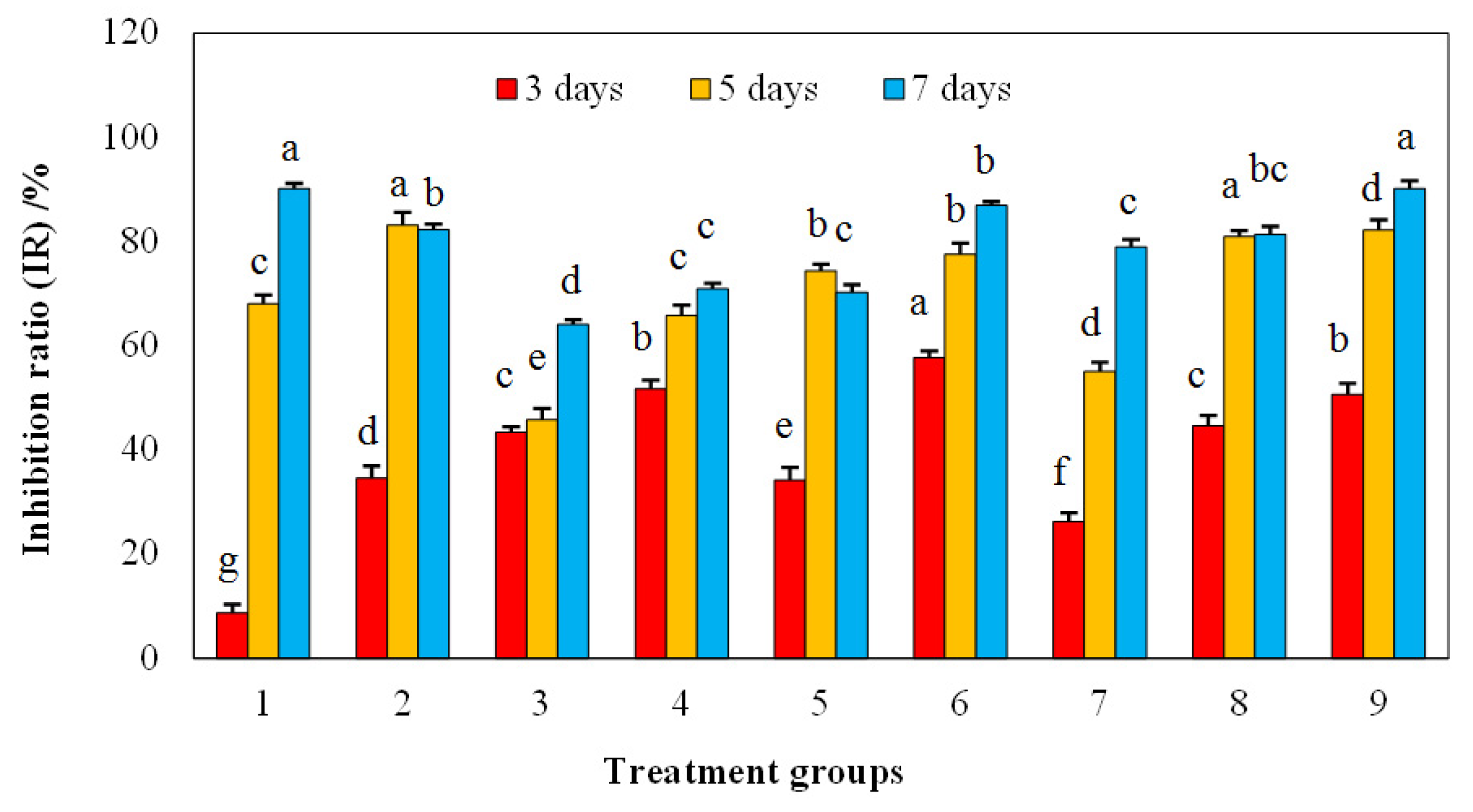
3.2. Inhibitory Effects of Mixed Allelochemicals on the Growth of Microcystis aeruginosa Cells
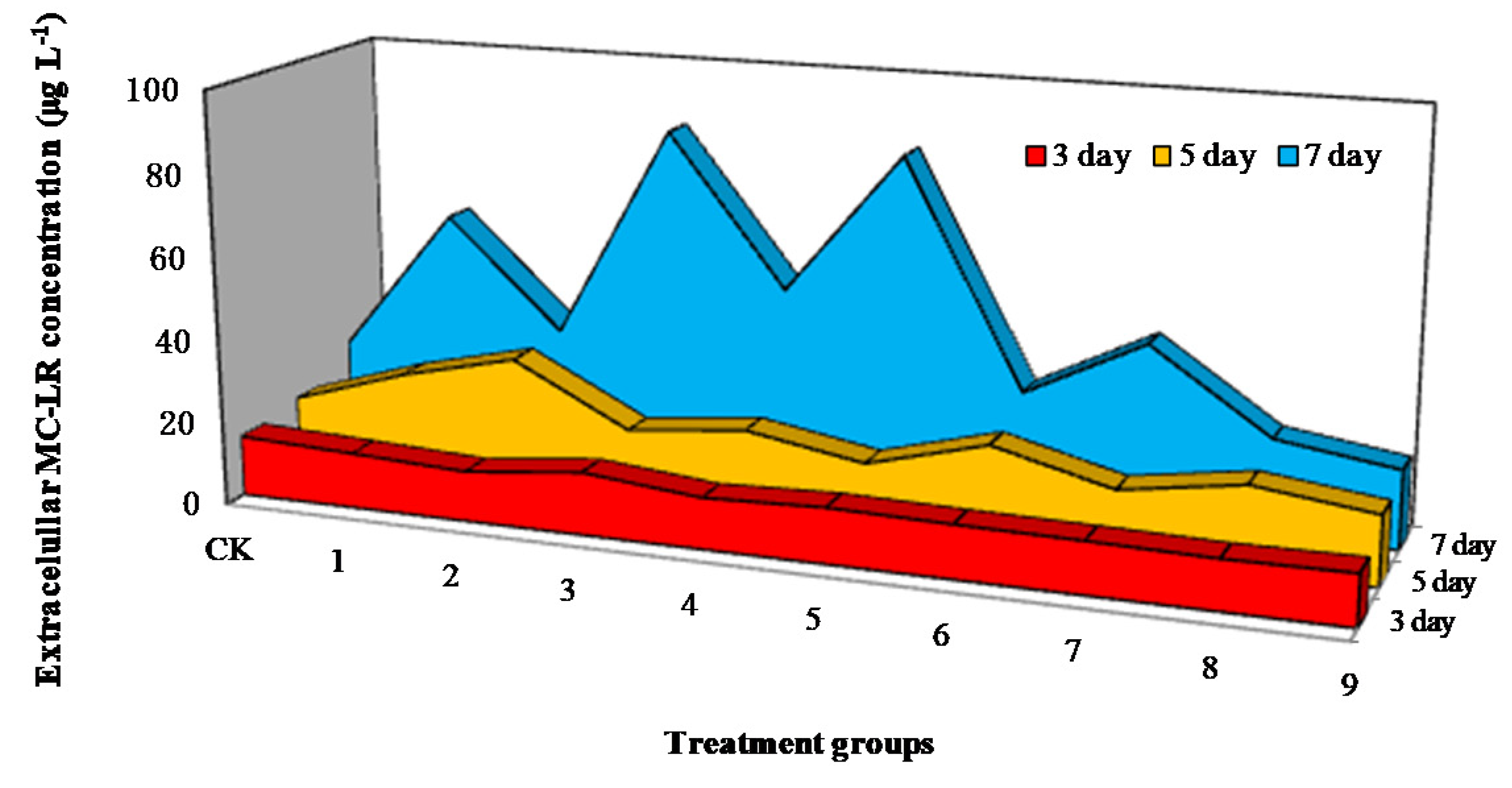
3.3. Influence of Mixed Allelochemicals on Extracellular MC-LR Content
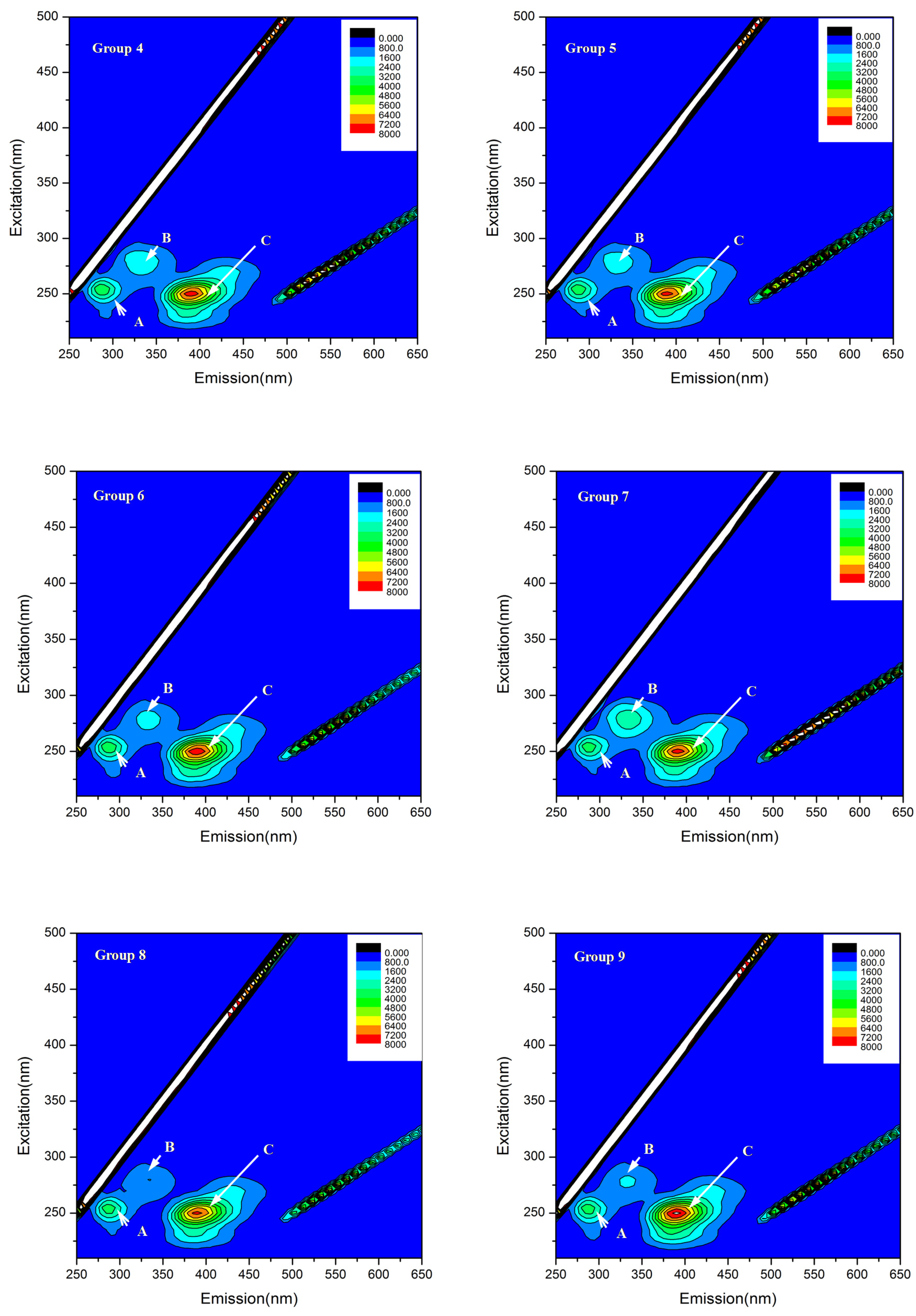
3.4. Influence of Mixed Allelochemicals on EPS Characterization
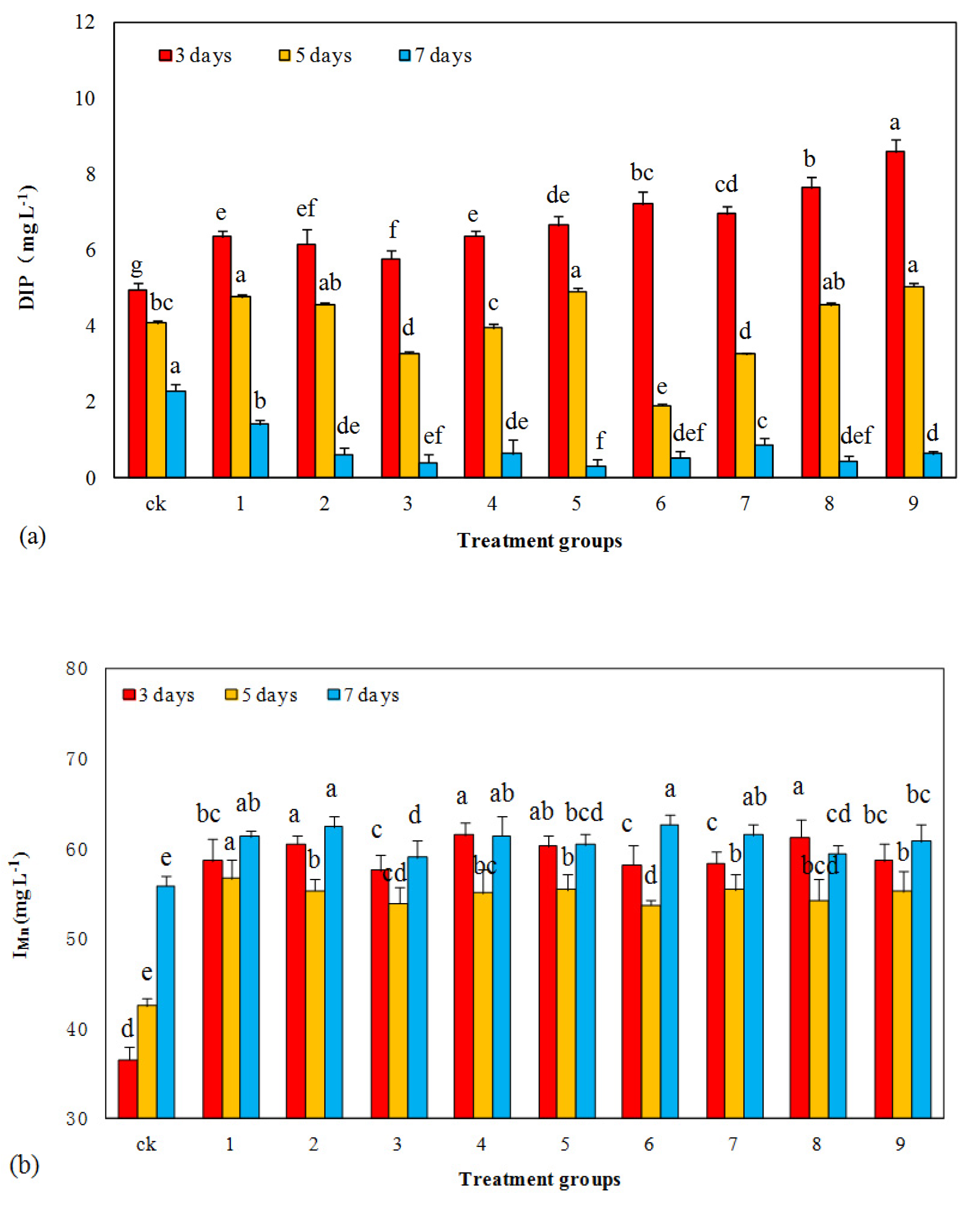
3.5. Influence of Mixed Allelochemicals on Imn and DIP Concentrations
4. Discussion
4.1. Contribution of the Allelochemicals to the Allelopathic Effect of Microcystis Aeruginosa
4.2. Compound Effects on Microcystis Aeruginosa Growth and Extracellular MC-LR
4.3. Possibility of EPS Variations of Microcystis Aeruginosa
4.4. Impacts on Water Quality
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, D.Y.; Chen, S.; Lu, J.; Liao, H.X. Corrigendum to ‘Study of the periphyton nutrient removal on eutrophic lake water quality’. Ecol. Eng. 2019, 130, 122–130. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Zamyadi, A.; Greenstein, K.E.; Glover, C.M.; Adams, C.; Rosenfeldt, E.; Wert, E.C. Impact of Hydrogen peroxide and copper sulfate on the delayed release of microcystin. Water 2020, 12, 1105. [Google Scholar] [CrossRef]
- Maršálek, B.; Maršálková, E.; Odehnalová, K.; Pochylý, F.; Rudolf, P.; Stahel, P.; Rahel, J.; Cech, J.; Fialová, S.; Zezulka, Š. Removal of Microcystis aeruginosa through the combined effect of plasma discharge and hydrodynamic cavitation. Water 2020, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hu, Y.D.; Zhu, M.; Yin, S.W. Integrated transcriptome and physiology analysis of Microcystis aeruginosa after exposure to copper sulfate. J. Oceanol. Limnol. 2020, 38, 102–113. [Google Scholar] [CrossRef]
- Techer, D.; Fontaine, P.; Personne, A.; Viot, S.; Thomas, M. Allelopathic potential and ecotoxicity evaluation of gallic and nonanoic acids to prevent cyanobacterial growth in lentic systems: A preliminary mesocosm study. Sci. Total Environ. 2016, 547, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Kaya, K.; Liu, Y.D.; Shen, Y.W.; Xiao, B.D.; Sano, T. Selective control of toxic Microcystis water blooms using lysine and malonic acid: An enclosure experiment. Environ. Toxicol. 2010, 20, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.X.; Acharya, K.; Hao, X.Y.; Li, S.Y. Isolation and identification of an antialgal compound from Artemisia annua and mechanisms of inhibitory effect on algae. Chemosphere 2012, 88, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.X.; Li, D.Y.; Hu, S.Z.; Wang, P.F.; Li, S.Y.; Li, Y.P.; Li, Y.; Acharya, K. Effects of artemisinin sustained-release granules on mixed alga growth and microcystins production and release. Environ. Sci. Pollut. Res. Int. 2015, 22, 18637–18644. [Google Scholar] [CrossRef] [PubMed]
- Nakai, S.; Yamada, S.; Hosomi, M. Anti-cyanobacterial fatty acids released from Myriophyllum spicatum. Hydrobiologia 2005, 543, 71–78. [Google Scholar] [CrossRef]
- Hehmann, A.; Watanabe, M.M.; Kaya, K. Killing of Microcystis by an amino acid: Results of laboratory and enclosure experiments. Verh. Int. Limnol. 2002, 28, 1147–1150. [Google Scholar] [CrossRef]
- Hu, X.; Liu, Y.G.; Zeng, G.M. Effects of D -menthol stress on the growth of and microcystin release by the freshwater cyanobacterium Microcystis aeruginosa FACHB-905. Chemosphere 2014, 113, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.Y.; Chen, Y.L.; Han, Y.Z.; Zhang, T.T. The inhibitory effect of lactic acid on Microcystis aeruginosa and its mechanisms. China Environ. Sci. 2016, 36, 1167–1172. [Google Scholar]
- Sebastián, E.S.; Beatríz, M.B.; Carlos, M.L.; Magdalena, S.J.; Macarena, P.; Marí, C.R.M. Microcystin accumulation and antioxidant responses in the freshwater clam Diplodon chilensis patagonicus upon subchronic exposure to toxic Microcystis aeruginosa. Ecotoxicol. Environ. Saf. 2011, 74, 1188–1194. [Google Scholar] [CrossRef]
- Boylan, J.; Morris, J. Limited Effects of Barley Straw on Algae and Zooplankton in a Midwestern Pond. Lake Reserv. Manag. 2003, 19, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Liu, Y.G.; Zeng, G.M.; Hu, X.J.; Wang, Y.Q.; Zeng, X.X. Effects of limonene stress on the growth of and microcystin release by the freshwater cyanobacterium Microcystis aeruginosa FACHB-905. Ecotoxicol. Environ. Saf. 2014, 105, 121–127. [Google Scholar] [CrossRef]
- Ni, L.X.; Jie, X.T.; Wang, P.F.; Li, S.Y.; Wang, G.X.; Li, Y.P.; Li, Y.; Acharya, K. Effect of linoleic acid sustained-release microspheres on Microcystis aeruginosa antioxidant enzymes activity and microcystins production and release. Chemosphere 2015, 121, 110–116. [Google Scholar] [CrossRef]
- Sheng, G.P.; Yu, H.Q.; Li, X.Y. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: A review. Biotechnol. Adv. 2010, 28, 882–894. [Google Scholar] [CrossRef]
- Xu, H.C.; Cai, H.Y.; Yu, G.H.; Jiang, H.L. Insights into extracellular polymeric substances of cyanobacterium Microcystis aeruginosa using fractionation procedure and parallel factor analysis. Water Res. 2013, 47, 2005–2014. [Google Scholar] [CrossRef]
- Liu, Z.; Cui, F.; Ma, H. The interaction between nitrobenzene and Microcystis aeruginosa and its potential to impact water quality. Chemosphere 2013, 92, 1201–1206. [Google Scholar] [CrossRef]
- García-Robledo, E.; Corzo, A. Effects of macroalgal blooms on carbon and nitrogen biogeochemical cycling in photoautotrophic sediments: An experimental mesocosm. Mar. Pollut. Bull. 2011, 62, 1550–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdemarsen, T.; Quintana, C.O.; Flindt, M.R. Organic N and P in eutrophic fjord sediments-rates of mineralization and consequences for internal nutrient loading. Biogeosciences 2015, 12, 1765–1779. [Google Scholar] [CrossRef] [Green Version]
- Rippka, R.; Deruelles, J.; Waterbury, J.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Jiang, H. Effects of cyanobacterial extracellular polymeric substances on the stability of ZnO nanoparticles in eutrophic shallow lakes. Environ. Pollut. 2015, 197, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, L.J.; Cheng, W.H.; Wu, D.L.; Pu, Y. Correlation Analysis of CODMn, BOD5 and CODcr in Surface Waters. Environ. Sci. Manag. (China) 2016, 41, 134–137. [Google Scholar] [CrossRef]
- Henriksen, T.; Snipes, W. Irradiation of dihydrothymine in frozen sulfuric acid solutions. Radiat. Res. 1970, 41, 439–449. [Google Scholar] [CrossRef]
- Ni, L.X.; Acharya, K.; Ren, G.X.; Li, S.Y.; Li, Y.P.; Li, Y. Preparation and characterization of anti-algal sustained-release granules and their inhibitory effects on algae. Chemosphere 2013, 91, 608–615. [Google Scholar] [CrossRef]
- Hong, Y.; Hu, H.Y. Research and application of aquatic plant allelopathy inhibition on algae. Chin. Sci. Bull. 2009, 54, 287–293. [Google Scholar] [CrossRef]
- Chang, X.X.; Eigemann, F.; Hilt, S. Do macrophytes support harmful cyanobacteria? Interactions with a green alga reverse the inhibiting effects of macrophyte allelochemicals on Microcystis aeruginosa. Harmful Algae 2002, 19, 76–84. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Su, Z.X.; Pu, Y.F.; Xiao, H.; Wang, C.H. Isolation, purification and identification of antialgal activity substances of ethyl acetate extracts from the submerged macrophytes Potamogeton crispus. Environ. Sci. 2015, 36, 3860–3865. [Google Scholar] [CrossRef]
- Streibig, J.C.; Olofsdotter, M. Joint action of phenolic acid mixtures and its significance in allelopathy research. Physiol. Plant. 2010, 114, 422–428. [Google Scholar] [CrossRef]
- Ni, L.X.; Ren, G.X.; Chen, S.J. Study on joint action of phenolic acids and unsaturated fatty acids to Microcystis aeruginosa. Environ. Chem. 2011, 30, 1428–1432. [Google Scholar]
- Xiao, X.; Chen, Y.X.; Liang, X.Q.; Lou, L.P.; Tang, X.J. Effects of Tibetan hulless barley on bloom-forming cyanobacterium (Microcystis aerugenosa) measured by different physiological and morphologic parameters. Chemosphere 2010, 81, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Sheng, G.P.; Xu, J.; Luo, H.W. Thermodynamic analysis on the binding of heavy metals onto extracellular polymeric substances (EPS) of activated sludge. Water Res. 2013, 47, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.Q.; Xiao, Y.; Zhu, L.; Wu, Z.X.; Liu, J.; Hu, C.L.; Song, L.R. The role of microcystins in maintaining colonies of bloom-forming Microcystis spp. Environ. Microbiol. 2012, 14, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.C.; He, P.J.; Wang, G.Z. Three-dimensional excitation emission matrix fluorescence spectroscopy and gel-permeating chromatography to characterize extracellular polymeric substances in aerobic granulation. Water Sci. Technol. 2010, 61, 2931–2942. [Google Scholar] [CrossRef]
- Yang, L.Y.; Wang, C.C.; Sun, X.; Guo, L.Y.; Xiao, L.; Song, X.J. Study on microbial nitrification and denitrification processes and influence factors in freshwater lakes. Water Resour. Prot. 2016, 32, 12–22. [Google Scholar]
- Sundback, K.; Miles, A.; Hulth, S.; Pihl, L. Importance of benthic nutrient regeneration during initiation of macroalgal blooms in shallow bays. Mar. Ecol. Prog. Ser. 2003, 246, 115–126. [Google Scholar] [CrossRef]
- Shapiro, J. Blue-green algae: Why they become dominant. Science 1973, 179, 382–384. [Google Scholar] [CrossRef]
- Peng, G.T.; Steven, W.W.; Lin, S.J.; Wang, X.R. Response of Microcystis aeruginosa FACHB-905 to different nutrient ratios and changes in phosphorus chemistry. J. Oceanol. Limnol. 2018, 36, 1040–1052. [Google Scholar] [CrossRef]
- Li, W.C.; Chen, K.; Wu, Q.L. Experimental studies on decomposition process of aquatic plant material from East Taihu Lake. J. Lake Sci. 2001, 13, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.J.; Qin, B.Q.; Zhu, G.W. Release of colloidal phosphorus, nitrogen and organic carbon in the course of dying and decomposing of cyanobacteria. China Environ. Sci. 2007, 27, 341–345. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group No. | Selected Chemicals | |||
|---|---|---|---|---|
| Artemisinin | Nonanoic Acid | Malonic Acid | Ethyl Acetate | |
| 1 | 1/9 EC50 | 1/9 EC50 | 1/9 EC50 | 1/9 EC50 |
| 2 | 1/9 EC50 | 1/6 EC50 | 1/6 EC50 | 1/6 EC50 |
| 3 | 1/9 EC50 | 1/3 EC50 | 1/3 EC50 | 1/3 EC50 |
| 4 | 1/6 EC50 | 1/9 EC50 | 1/6 EC50 | 1/3 EC50 |
| 5 | 1/6 EC50 | 1/6 EC50 | 1/3 EC50 | 1/9 EC50 |
| 6 | 1/6 EC50 | 1/3 EC50 | 1/9 EC50 | 1/6 EC50 |
| 7 | 1/3 EC50 | 1/9 EC50 | 1/3 EC50 | 1/6 EC50 |
| 8 | 1/3 EC50 | 1/6 EC50 | 1/9 EC50 | 1/3 EC50 |
| 9 | 1/3 EC50 | 1/3 EC50 | 1/6 EC50 | 1/9 EC50 |
| Group | Artemisinin | Nonanoic Acid | Malonic Acid | ||||||
| ± S | F | P | ± S | F | P | ± S | F | P | |
| 0 mg L−1 | 17.53 ± 0.09 | 1291.949 | P < 0.001 | 17.53 ± 0.09 | 203.552 | P < 0.001 | 8.64 ± 0.19 | 120.959 | P < 0.001 |
| 5 mg L−1 | 12.56 ± 0.37 | 12.13 ± 0.88 | 9.57 ± 0.37 | ||||||
| 10 mg L−1 | 9.77 ± 0.22 | 9.24 ± 0.72 | 9.25 ± 0.72 | ||||||
| 20 mg L−1 | 8.71 ± 0.41 | 7.41 ± 0.41 | 9.16 ± 0.84 | ||||||
| 40 mg L−1 | 6.17 ± 0.06 | 5.57 ± 0.51 | 6.27 ± 0.14 | ||||||
| 60 mg L−1 | 4.32 ± 0.09 | 4.91 ± 0.44 | 4.33 ± 0.09 | ||||||
| 80 mg L−1 | 4.34 ± 0.15 | 5.11 ± 0.58 | 3.35 ± 0.15 | ||||||
| 120 mg L−1 | 4.87 ± 0.10 | 5.81 ± 0.20 | 3.19 ± 0.29 | ||||||
| Group | D-menthol | Lactic Acid | Ethyl Acetate | ||||||
| ± S | F | P | ± S | F | P | ± S | F | P | |
| 0 mg L−1 | 8.64 ± 0.19 | 120.959 | P < 0.001 | 17.53 ± 0.09 | 319.506 | P < 0.001 | 17.53 ± 0.09 | 393.855 | P < 0.001 |
| 5 mg L−1 | 9.65 ± 0.37 | 16.21 ± 0.23 | 21.03 ± 1.14 | ||||||
| 10 mg L−1 | 8.65 ± 0.27 | 15.70 ± 0.23 | 19.79 ± 0.23 | ||||||
| 20 mg L−1 | 7.70 ± 0.72 | 14.38 ± 0.45 | 11.67 ± 0.69 | ||||||
| 40 mg L−1 | 6.60 ± 0.32 | 15.80 ± 0.69 | 7.38 ± 0.72 | ||||||
| 60 mg L−1 | 6.59 ± 0.58 | 16.48 ± 0.29 | 6.35 ± 0.10 | ||||||
| 80 mg L−1 | 5.08 ± 0.53 | 5.18 ± 0.85 | 6.26 ± 0.22 | ||||||
| 120 mg L−1 | 4.54 ± 0.44 | 5.58 ± 0.53 | 5.42 ± 0.46 | ||||||
| Chemicals | Regression Equation | 95% Confidence Limits | EC50 |
|---|---|---|---|
| Probit (Y) = b × logarithm (X) + a | |||
| Artemisinin | Y = 0.016X − 0.384 | b 0.016~0.016 | 23.66 |
| a −0.385~−0.383 | |||
| Nonanoic acid | Y = 0.029X − 0.537 | b 0.029~0.029 | 18.81 |
| a −0.539~−0.535 | |||
| Malonic acid | Y = 0.017X − 1.262 | b 0.017~0.017 | 73.8 |
| a −1.265~−1.258 | |||
| D-menthol | Y = 0.018X − 2.044 | b 0.018~0.018 | 115.72 |
| a −2.047~−2.041 | |||
| lactic acid | Y = 0.034X − 2.943 | b 0.034~0.034 | 86.21 |
| a −2.944~−2.942 | |||
| Ethyl acetate | Y = 0.013X − 0.49 | b 0.013~0.013 | 38.26 |
| a −0.492~−0.488 |
| Group No. | Peaks | Ex/Em | Intensity | Corresponding Peaks |
|---|---|---|---|---|
| Control | A | 255/285 | 3890 | Tyrosine and protein-like compounds |
| B | 285/315 | 822 | Tryptophan containing protein-like compounds | |
| C | 250/390 | 7397 | Fulvic acid-like compounds | |
| 1 | A | 255/285 | 3825 | Tyrosine and protein-like compounds |
| B | 280/335 | 1089 | Tryptophan | |
| C | 250/390 | 7489 | Fulvic acid-like compounds | |
| 2 | A | 255/285 | 3888 | Tyrosine and protein-like compounds |
| B | 280/335 | 1229 | Tryptophan | |
| C | 250/390 | 7694 | Fulvic acid-like compounds | |
| 3 | A | 255/285 | 3963 | Tyrosine and protein-like compounds |
| B | 285/320 | 1149 | Tryptophan containing protein-like compounds | |
| C | 250/390 | 8291 | Fulvic acid-like compounds | |
| 4 | A | 255/285 | 3835 | Tyrosine and protein-like compounds |
| B | 280/335 | 2226 | Tryptophan | |
| C | 250/390 | 7765 | Fulvic acid-like compounds | |
| 5 | A | 255/285 | 3832 | Tyrosine and protein-like compounds |
| B | 280/330 | 2506 | Tryptophan | |
| C | 250/390 | 8002 | Fulvic acid-like compounds | |
| 6 | A | 255/285 | 3866 | Tyrosine and protein-like compounds |
| B | 280/335 | 2971 | Tryptophan | |
| C | 250/390 | 7798 | Fulvic acid-like compounds | |
| 7 | A | 255/285 | 3715 | Tyrosine and protein-like compounds |
| B | 280/335 | 3004 | Tryptophan | |
| C | 250/390 | 7855 | Fulvic acid-like compounds | |
| 8 | A | 255/285 | 3876 | Tyrosine and protein-like compounds |
| B | 280/335 | 1837 | Tryptophan | |
| C | 250/390 | 8339 | Fulvic acid-like compounds | |
| 9 | A | 255/285 | 3869 | Tyrosine and protein-like compounds |
| B | 280/335 | 2015 | Tryptophan | |
| C | 250/390 | 8465 | Fulvic acid-like compounds |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, P.; Wang, C.; Wang, P.; Gan, X.; Wang, X.; Yang, C. Effects of Mixed Allelochemicals on the Growth of Microcystis aeruginosa, Microcystin Production, Extracellular Polymeric Substances, and Water Quality. Water 2020, 12, 1861. https://doi.org/10.3390/w12071861
Ouyang P, Wang C, Wang P, Gan X, Wang X, Yang C. Effects of Mixed Allelochemicals on the Growth of Microcystis aeruginosa, Microcystin Production, Extracellular Polymeric Substances, and Water Quality. Water. 2020; 12(7):1861. https://doi.org/10.3390/w12071861
Chicago/Turabian StyleOuyang, Ping, Chao Wang, Peifang Wang, Xiaorong Gan, Xun Wang, and Chaohui Yang. 2020. "Effects of Mixed Allelochemicals on the Growth of Microcystis aeruginosa, Microcystin Production, Extracellular Polymeric Substances, and Water Quality" Water 12, no. 7: 1861. https://doi.org/10.3390/w12071861