Trajectories of Sediment-Water Interactions in Reservoirs as a Result of Temperature and Oxygen Conditions


UFZ-Helmholtz Centre for Environmental Research, Department Lake Research, Brueckstr. 3a, D-39114 Magdeburg, Germany
*
Author to whom correspondence should be addressed.
Water 2020, 12(4), 1065; https://doi.org/10.3390/w12041065
Submission received: 11 February 2020
/
Revised: 4 April 2020
/
Accepted: 7 April 2020
/
Published: 9 April 2020
(This article belongs to the Section Water Quality and Contamination)
Abstract
:Temperate lakes/reservoirs are warming; this can influence the benthic release of nutrients. They undergo seasonal changes resulting in an array of temperature and oxygen conditions; oxic-low, oxic-high, anoxic-low, and anoxic-high temperature. We sought to understand the interaction of temperature and oxygen conditions on benthic solutes exchange through a two-factorial sediment core incubation experiment by varying either temperature or oxygen conditions of sediment cores from an oligotrophic and eutrophic reservoir. Temperature and oxygen conditions are both important for nutrient release; however, they influence solutes differently; differences in the fluxes of the treatments were explained more by temperature for P, DOC and N, while for Fe, Mn and SO42−, differences were explained more by oxygen conditions. The combination of strongly reducing conditions (due to anoxia) and high temperature (20 °C) led to a significant increase in nutrients concentrations in the overlying water. Under these conditions, SRP flux was 0.04 and 0.5 mmol m−2 d−1; ammonium was 0.9 and 5.6 mmol m−2 d−1 for the oligotrophic and eutrophic reservoir, respectively. We observed a synergistic interaction between temperature and oxygen conditions which resulted in release of solutes from sediments. An increase in nutrients release under increasing temperatures is more likely and so are algal blooms.
1. Introduction
Temperate lakes and reservoirs undergo seasonal changes in temperature and sometimes oxygen conditions, which can affect benthic nutrients release. They mix during cold seasons and their sediments are normally exposed to low temperature and oxic conditions. Over the warm season, these reservoirs can stratify or remain mixed and the sediments may remain cold or experience substantial warming. The redox conditions may remain high as long as oxygen is available at sufficient quantities or become negative when oxygen is depleted. The distinct effects of temperature and oxygen conditions on sediment–water interactions are known but their interaction effects are not well quantified. This is important because temperature and oxygen conditions are interlinked by indirect effects, e.g., does a higher temperature accelerate oxygen depletion and thus support the occurrence of low redox potentials (see below). Such interactions can be highly relevant in lake management as ecological status and water quality largely depend on the nutrients concentrations and biogeochemical processes. The sediment is an important compartment for biogeochemical processes, where key processes take place, i.e., release of phosphorus, carbon, and metals [1,2,3], oxygen depletion [4], and nitrogen removal by denitrification [5]. Benthic nutrients release or internal loading is a major source of nutrients in many water bodies [6]. Hence, experimental testing of temperature-oxygen interactions on benthic nutrient release is of fundamental importance for water quality management.
Temperature is critical factor regulating benthic biogeochemical processes in lakes/reservoirs [7]. Hypolimnion temperatures in deep temperate lakes/reservoirs are usually low (4–6 °C) and constant or only slightly changing over the course of the year [3]. Temperature profile data in stratified temperate lakes/reservoirs with moderate depth, however, can also show hypolimnetic temperatures greater than 10 °C [8]. In addition, hypolimnetic water temperatures are increasing in many water bodies) due to climate warming as observed in Lake Lugano [9] and several others [10]. These observations raise questions about temperature effects on mobilization of nutrients, dissolved organic carbon (DOC) and metals from sediments and the associated implications for lake ecosystem functioning.
Rising temperatures at the sediment–water interface intensify the release of phosphorus from sediments [11] potentially promoting eutrophication and the occurrence of cyanobacterial blooms [12,13]. At the start of spring, as the lake temperature start to increase, so does all temperature dependent process like primary production and mineralization of organic carbon in the sediment. The increase in water temperature accelerates organic matter mineralization and other processes hence the uptake of oxygen at the sediment–water interface [14] and can induce hypoxic or anoxic conditions. If the P stored in the sediment is redox dependent this leads to release of P into the water column thereby providing nutrients for algal growth from both processes of organic matter mineralization and redox dependent release explained below. Another negative effect of warming on oxygen concentration is mediated by the extension of stratification duration in a warmer climate [15]. Longer stratification duration prolongs oxygen depletion and can induce anoxia and further negative effects on water quality, as shown in Lake Zürich [16]. Temperature-driven hypoxia is now a threat in several lakes, even in those that previously were well oxygenated [17]. Furthermore, modelling of climate change effects already shows increasing risk of anoxia in lakes [18] and hence, the recovery of lakes from eutrophication will become more difficult [19].
Redox condition is another major determinant of sediment–water interactions [20,21], which affects the dominant biogeochemical redox reactions and solubility of some metals and ions. At low redox potential, which occurs at low oxygen and nitrate conditions, iron and manganese are reduced and released into the water [2,22]. This reductive dissolution of Fe causes the release of P and DOC [2,3], making the oxygen conditions of a lake’s hypolimnion a key issue in lake management.
The effects of temperature and oxygen conditions on benthic solute fluxes have been extensively studied, but only separately [7,11,23,24,25]. Furthermore, recent studies on sediment–water interactions displayed a selective focus on carbon-related topics such as methane and carbon dioxide fluxes or the mineralization of organic carbon. Hence, existing studies rarely address a complete assessment of benthic fluxes, including nutrients, carbon, metals and other solutes, and are not covering the entire trophic state spectrum of lakes/reservoirs [26,27]. We hypothesize that the interaction of temperature and oxygen state strongly affect sediment–water interactions and focus on quantifying these effects in an experimental setting.
We designed this study to achieve a quantitative assessment of sediment–water interactions at different conditions of temperature and oxygen by using an experimental approach. We chose two reservoirs that differ strongly in terms of morphology and trophic state: the oligotrophic Rappbode Reservoir (Oligo-R) and the eutrophic Bautzen Reservoir (Eutro-B). These two water bodies cover the full range of trophic state and hence, were selected to reflect contrasting sediment characteristics and nutrient loading regimes. In both reservoirs, sediments experience very similar conditions during the cold season (i.e., cold and oxic) but are exposed to all combinations of warm versus cold and oxic versus anoxic conditions during late summer (Figure 1). The following four combinations were investigated; (1) oxic-low temperature; (2) anoxic-low temperature; (3) oxic-high temperature; and (4) anoxic-high temperature. We hypothesize that the combination of warming and anoxia enhance the exchange of nutrients and metals at the sediment–water interface. We also hypothesize that the interaction of warming and anoxia is synergistic and intensify solute release rates disproportionally. We tested these hypotheses through a fully crossed two-factorial sediment core incubation experiment by varying either temperature or oxygen conditions of the sediment. Our assessment included solute fluxes of phosphorus, nitrate, ammonia, iron, manganese, dissolved organic carbon (DOC), oxygen and sulphate in order to accomplish a deeper understanding of benthic solutes exchange under different environmental conditions and to provide insight into the consequences of climate warming.
2. Materials and Methods
2.1. Study Sites
Sediments were extracted from Rappbode reservoir (Oligo-R) and from Bautzen reservoir (Eutro-B) in Germany; the reservoir characteristics are summarized in Table 1. Rappbode reservoir is located in the eastern Harz Mountains (51.74° N, 10.89° E) and is the largest drinking water reservoir in Germany [28]. It is an oligotrophic reservoir with a dimictic mixing pattern. Rappbode experiences water level fluctuations of up to 15 m during the year [29]. Bautzen is located in the upper catchment of the River Spree directly downstream of the city of Bautzen (51.218° N, 14.466° E). The primary use of the reservoir is maintaining a minimum summer water discharge in the lower River Spree in order to supply water to downstream wetlands and thermal power plants. The dimictic reservoir can display a polymictic character in some years due to its exposed location in a windy area and the weak stratification [30]. The reservoir operation often includes high water withdrawal during summer which usually goes along with major drops in water level. For simplicity the terms Oligo-R and Eutro-B is mostly used in the text to refer to Rappbode and Bautzen reservoirs, respectively.
2.2. Sediment and Water Sampling
Sampling was carried out in September 2015 at the Oligo-R (17 m depth) and in March 2017 at the Eutro-B (13 m depth); the sediment–water interface was oxic in both locations. Water samples were collected in the hypolimnion using a standard 2 litre water sampler (Hydrobios, Kiel, Germany) and analyzed for various parameters (see water analyses section below). Profiles of temperature, oxygen, pH, turbidity, and chlorophyll a were measured using the EXO2 (YSI, Yellow Springs, OH, USA) multi-parameter sonde equipped with corresponding calibrated sensors.
Undisturbed sediment cores with a mean overlying water height of 26 ± 7 cm (1.5 L) for Oligo-R (12 cores) and 20 ± 4 cm (1.2 L) for Eutro-B (16 cores) were retrieved from each reservoir (Eutro-B at 51.219° N, 14.465° E and Oligo-R at 51.736° N, 10.871° E) by using a modified Kajak gravity corer (UWITEC, Mondsee, Austria) and plastic tubes of 60 cm length with an inner diameter of 9 cm. The cores were closed (at the bottom and at the top) immediately after retrieving them. At the shore line the top cover was replaced with a special air tight lid which allowed continuous stirring of the overlying water phase to ensure homogeneity within the water phase. The lid also has ports for sample extraction, pH and redox electrodes, oxygen sensors, and bubbling (to maintain oxic or anoxic conditions, see below). Details of the structure and functioning of this specialized lid can be found in [37].
2.3. Sediment Incubation
In the laboratory, the cores were incubated in climate chambers at two different temperatures, 6 °C and 20 °C (±1 °C accuracy), respectively. We applied a 2 × 2-factorial design with 3 and 4 replicates cores for Oligo-R and Eutro-B, respectively. The factors were temperature and oxygen conditions, each with two levels. The resulting four treatment combinations were; (i) oxic-6 °C, (ii) oxic-20 °C, (iii) anoxic-6 °C, and (iv) anoxic-20 °C. All cores were wrapped with an aluminum foil to avoid light and thereafter anoxic-6 °C and anoxic-20 °C were made anoxic by bubbling with a mixture of nitrogen and carbon dioxide (99.96% N2/0.04% CO2) whereas the remaining oxic-6 °C and at oxic-20 °C cores were kept oxic by gently bubbling with air [37]. The oxic and anoxic conditions and temperature were monitored by a FireStingO2 optical oxygen meter with optical oxygen (O2) sensors and a temperature sensor (Pyro Science, Aachen, Germany). To determine the oxygen consumption rate in the oxic cores, oxygen was measured continuously for 3 h; this was only done for Eutro-B incubation. Cores were left to acclimatize to the temperature and oxygen condition treatments for 2 days prior to sampling of the overlying water. We incubated the sediment cores for 14 and 19 days for Eutro-B and Oligo-R respectively. During the incubation period the overlying water in each core was sampled 5 and 7 times for Eutro-B and Oligo-R, respectively, using a syringe. The incubation samples were analyzed for DP, SRP, DOC, NH4+-N, SO42−, Fe (dissolved), and Mn (dissolved).
Redox potential and pH were measured in the overlying water at the time of sampling with a multi-parameter meter fitted with conventional pH and redox potential electrodes (WTW Multi3430, Germany). Redox potential was only measured in Eutro-B. To convert the measured oxidation-reduction potential (ORP) to the standard hydrogen electrode potential a correction factor of +221 mV at 5 °C was applied, based on the ORP electrode operating manual. The sampled water volume (about 60 mL) was replaced by water from the hypolimnion of both reservoirs taken during sediment coring (approx. 40 L). Replacement water solutes concentrations were monitored and accounted for in the calculations of actual concentration in the cores. For oxic cores, oxic hypolimnion water was used as replacement water while for anoxic cores; hypolimnion water that had been flushed with N2/CO2 gas was used. From this replacement water, 4 bottles (controls) of 1 L volume were filled and incubated at the same conditions as the cores to account for changes occurring in the water that are not influenced by the sediment. The bottles were sampled 4 times during the incubation, i.e., every 2nd sampling of the cores. This was done because changes in concentration of solutes in controls were minimal in previous studies [3,37].
2.4. Water Analyses
Water samples from the reservoirs and water samples from the sediment incubation (selected parameters, see above) were analyzed for the following chemical parameters: dissolved phosphorus (DP), total phosphorus (TP), soluble reactive phosphorus (SRP), dissolved organic carbon (DOC), nitrate (NO3−-N), ammonium (NH4+-N), sulphate (SO42−), total dissolved iron (Fe), and total dissolved manganese (Mn). All samples were filtered by 0.2 µm syringe filter (Sartorius, Göttingen, Germany) prior to analyses. Dissolved phosphorus, SRP, NO3−-N, NH4+-N and TP (after digestion) were analyzed photometrically (Skalar, Breda, The Netherlands), DOC was measured with a C-analyzer (Dimatec, Essen, Germany), Fe and Mn were determined by ICP-OES (Perkin Elmer, Überlingen, Germany), and SO42− were analyzed by ion-chromatography (Dionex). Details of the analytical methods used for the chemical analysis are found in [35].
2.5. Sediment Characterization
For general sediment characterization (pH, redox potential, water content, loss on ignition, C, N and S), porewater extraction and P-fractionation analysis; three extra sediments cores were extracted from Bautzen at the time of sediment sampling. For Rappbode an extra core was extracted for general sediment characterization; for porewater extraction and P-fractionation we used the incubation sediment cores from the oxic-low temperature treatment at the end of incubation. Sediment cores for porewater extraction and P fractionation were sectioned into the following layers; 0–1, 1–2, 2–3, 3–4, 4–5, 5–6, 6–8, 8–10 cm, while those for elemental X-ray fluorescence analysis were sectioned in 2 cm layers. The sediments for general sediment characterization were measured for redox potential and pH in the field using multi-parameter meter fitted with conventional pH and redox potential electrodes (WTW Multi3430, Weilheim, Germany). Porewater was extracted by centrifugation (3500 rpm at 6 °C) and the samples were filtered using 0.45 µm syringe filter (Sartorius, Göttingen, Germany). Porewater samples were analyzed for SRP, Fe (dissolved), Mn (dissolved) SO42−, NH4+-N, and DOC as described in the previous section.
Water content was determined by drying sediment samples at 105 °C, taking note of sediment weight prior to and after drying. Loss on ignition was determined as the difference in weight of sediment samples prior and after combustion in an oven at 550 °C for 2 h. Total carbon (TC), total organic carbon (TOC, after removal of inorganic carbon by acidification), and total nitrogen (TN) were determined by a CN analyzer vario EL cube (Elementar Analysensysteme GmbH, Hanau, Germany). Total phosphorus (TP) was analyzed photometrically (Skalar, The Netherlands) after combustion at 550 °C for 2 h and boiling in 1 N HCl for 15 min. Iron and aluminum in sediments were determined by wavelength dispersive X-ray fluorescence (XRF; S4 Pioneer Bruker-AXS) according to the method described by [38].
2.6. Phosphorus Fractionation
Phosphorus fractionation was carried using a sequential extraction method of [39] modified by [40], which separates the following P fractions:
- Easily available, dissolved porewater P (NH4Cl-P) extracted by 1 M NH4Cl (ammonium chloride).
- Fe bound, redox-sensitive (BD-P) P extracted by 0.11 M NaHCO3 (sodium hydrogen carbonate)/0.11 M Na2S2O4 (sodium dithionite).
- Al-associated (NaOH-P) extracted by 1 M NaOH (sodium hydroxide), this P fraction is subdivided into NaOH-SRP (OH-exchangeable P at high pH) and NaOH-NRP (organic bound P).
- P bound by carbonates and apatite (HCl-P) extracted by 0.5 M HCl (hydrochloric acid).
- Refractory P (residual-P) determined by digestion of the remaining sediment with persulphate (K2S2O8) and sulphiric acid (H2SO4) at 110 °C for 30 min.
2.7. Calculation of Solute Release Rates
To calculate solutes fluxes, concentrations were corrected by including the amount and concentration of the water used to replace sample volumes. A linear relationship was assumed for the change in concentration over time. The calculation of the solute flux was performed according to [41]:
where = solute release rate (mg m−2 day−1), = solute concentration at time t (mg L−1), = solute concentration at t = 0 (mg L−1), = Volume of the water column (L), = surface area of the sediment (m2), = duration of the experiment (given in days).
Equation (1) describes a linear relationship of the solute flux with time. Since all the experiments were done in triplicate, the slope of the regression line from the linear fit was used instead of the term . A positive value indicates solute release (out-flux) from sediment, a negative value indicates solute uptake (in-flux) by the sediment.
2.8. Statistics
All statistics were done in R [42] under version R 3.5.1. The experimental results were analyzed by analysis of variance (ANOVA) by testing for effects from the two treatments: oxygen conditions (oxic vs. anoxic conditions) and temperature (low vs. high) as well as a potential interaction. Since the experimental design was sometimes unbalanced (unequal number of replicates for the treatments) we adopted type-II ANOVA by using the method Anova () from the package car that is able to calculate correct p-values from unbalanced designs. Bautzen oxic-low temperature treatment had only 3 replicates because 1 replicate got lost, while all the other treatments had 4 replicates each. We decided to analyze results for each reservoir separately because the studied variables did not react homogeneously in the two reservoirs. Please note that a significant interaction means that both main factors are not independent from each other, i.e., they are not interacting in an additive way but rather synergistically. This is the case, for example, whenever the effect of temperature is not the same within each oxygen treatment. We also computed the overall coefficient of determination (R2) for the underlying linear model and attributed the explained variance to the different model terms by calculating partial R2 using methods from [43]. This was achieved by using the method calc. relimp () from the package relaimpo.
3. Results
3.1. Water, Sediment and Porewater Characteristics
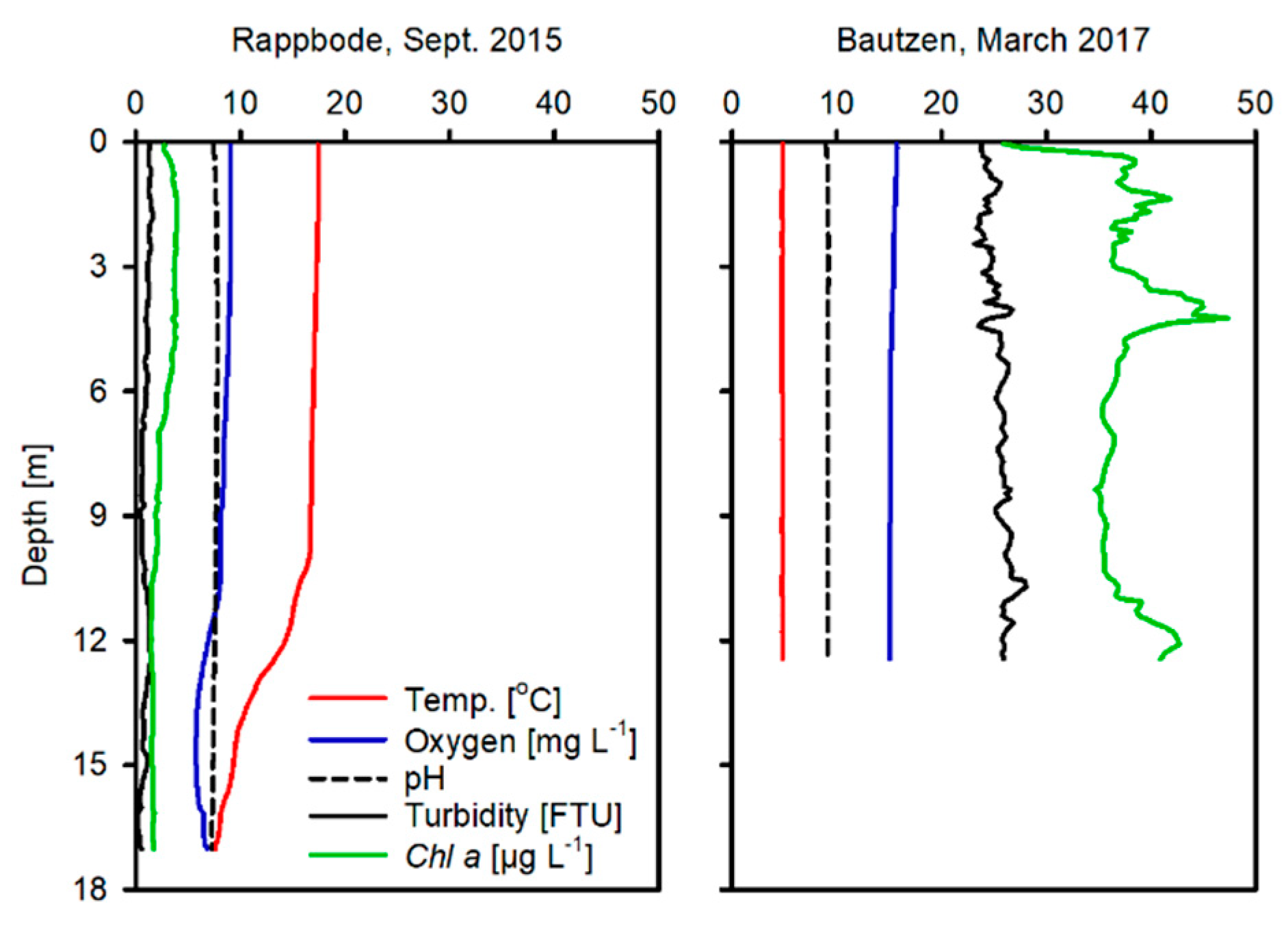
Water quality profiles taken at the reservoirs prior to sampling show that both Rappbode (Oligo-R) and Bautzen reservoir (Eutro-B) were oxic with Oligo-R having pronounced temperature stratification (Figure 2). Chlorophyll and turbidity was very high in Eutro-B in comparison to Oligo-R (Figure 2). Oligo-R pH (Figure 2) was circum-neutral (7.3) while that of Eutro-B was more alkaline (9.1).
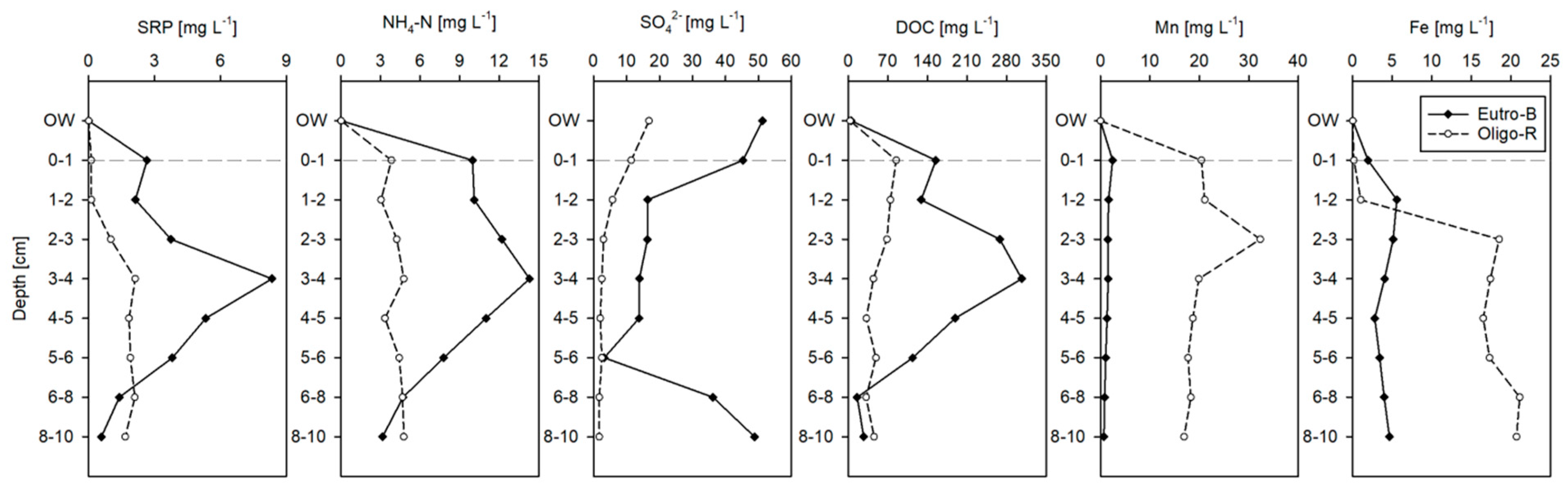
Solute concentrations in the porewater were generally higher than in the hypolimnion with the exception of SO42− (Figure 3). Generally, Eutro-B had a higher solute concentration than Oligo-R in both overlying water and porewater, with the exception of Mn and Fe that were higher in Oligo-R. The solute concentration was more variable with depth in Eutro-B in comparison with Oligo-R. In Eutro-B there is a concentration peak in the 3–4 cm layer for SRP, NH4+-N and SO42−. Both reservoirs had similar nutrient concentrations (SRP and ammonia) in the 6–8 and 8–10 cm layers (Figure 3). Total phosphorus (TP) in the overlying (hypolimnion) water was 0.008 and 0.073 mg L−1, while NO3−-N was 1.31 and 2.96 mg L−1 in Oligo-R and Eutro-B, respectively.
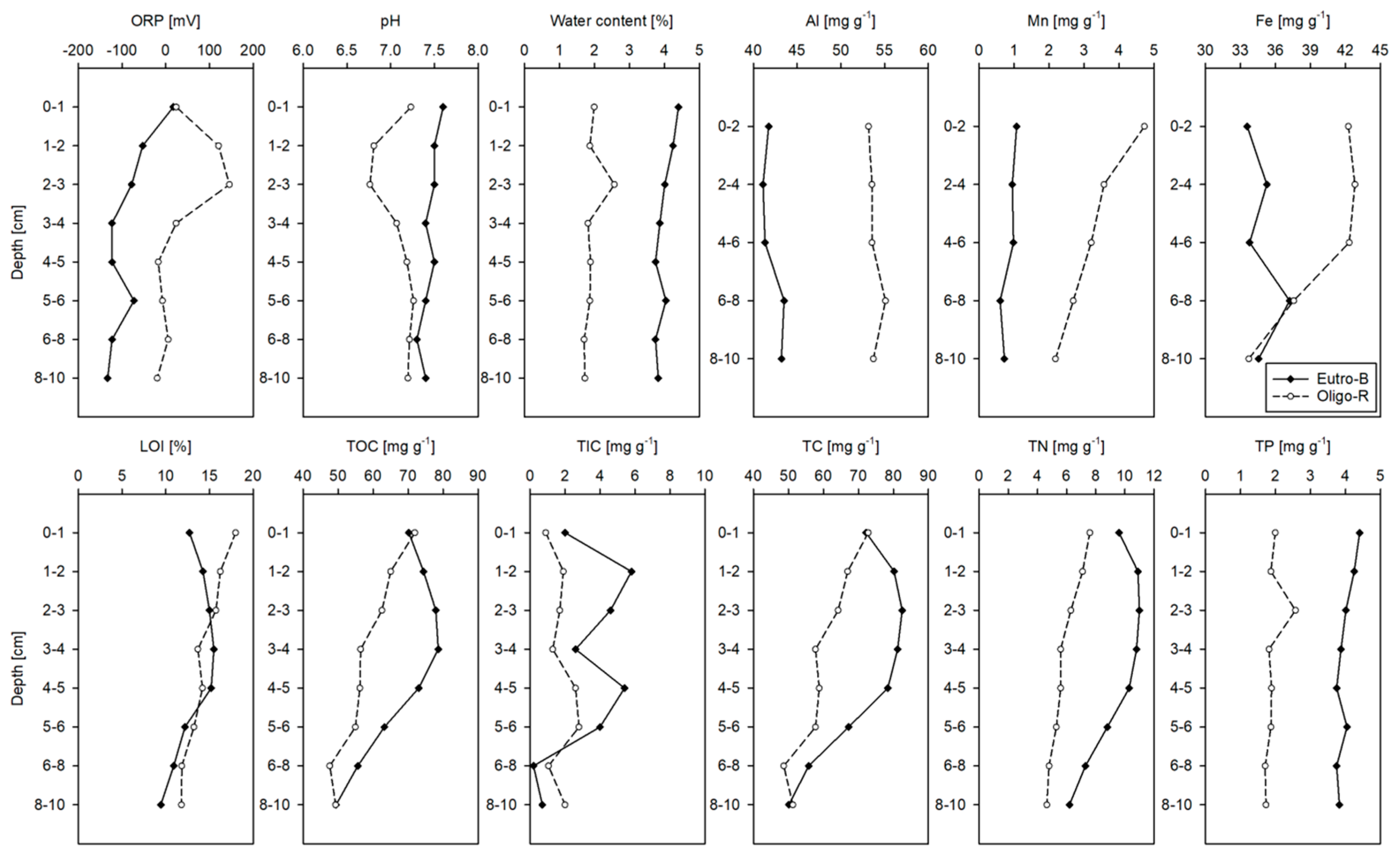
The redox potential profile for Eutro-B was lower than for Oligo-R especially for the 1–2 and 2–3 cm depth (Figure 4). Sediment pH profiles for both reservoirs remained in the neutral range. TN and TP were higher in Eutro-B than Oligo-R, with TP in Eutro-B twice that in Oligo-R (Figure 4), while the metals (Fe, Mn, and Al) had an opposite pattern. Water content, LOI, TC, TOC, and TN in both reservoirs decreased with depth.
3.2. Sediment Incubation Conditions
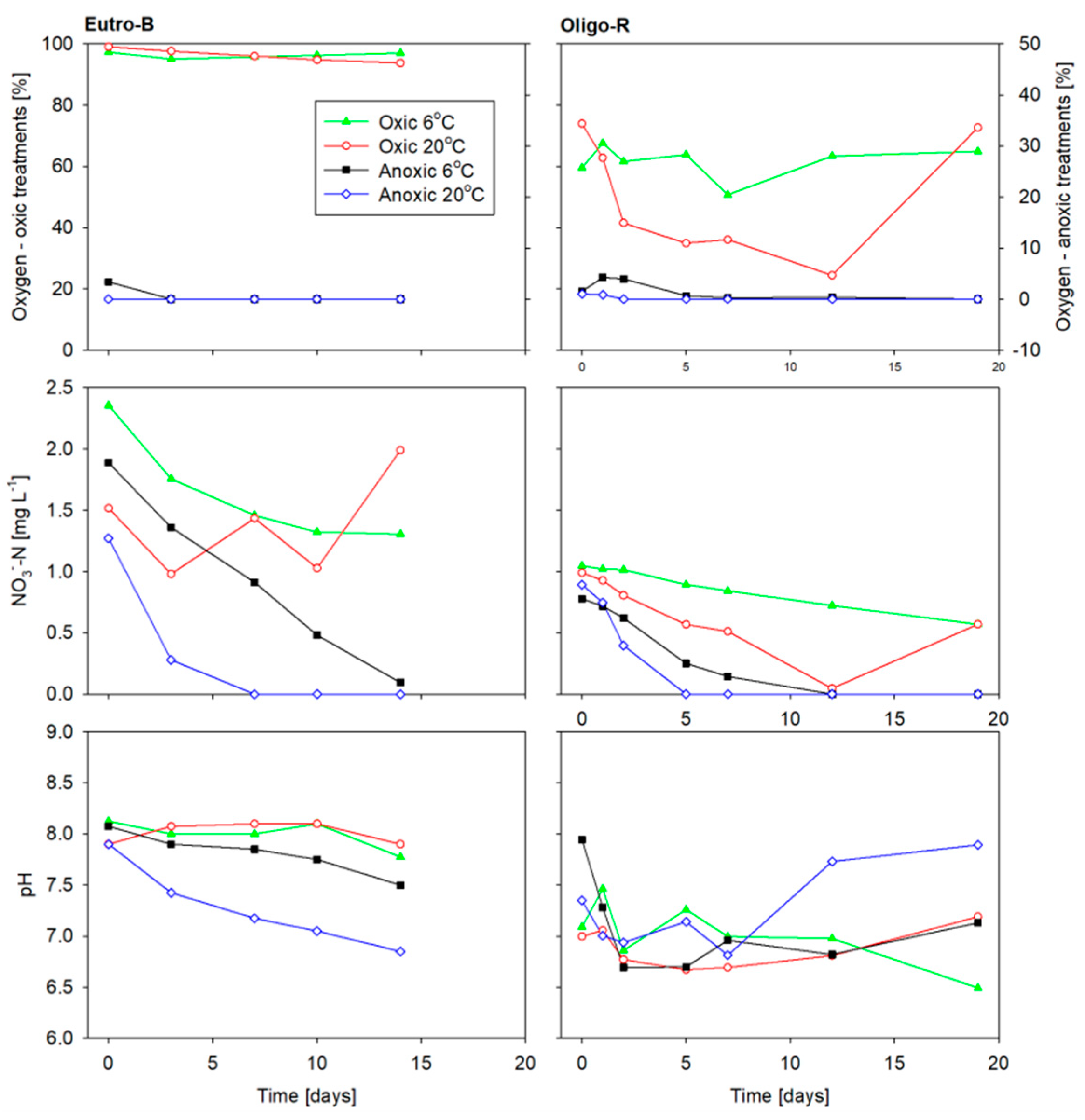
Mean oxygen saturation in the Oligo-R oxic treatments was 63% and 53% for the low temperature and high temperature treatments, respectively, due to problems with air bubbling mechanism. Eutro-B mean oxygen saturation was above 96% for all oxic treatments (Figure 5). All anoxic treatments from both Oligo-R and Eutro-B had no oxygen (Figure 5). Nitrate decreased below limit of quantification (0.042 mg L−1) in the anoxic-high temperature treatment after seven days in Eutro B and 5 days in Oligo-R (Figure 5). Moreover, in the anoxic-low temperature treatments, nitrate depletion took place but at slower rates than at the high temperature (Figure 5). In the oxic treatments nitrate decreased gradually during the incubation period, although also shorter periods of nitrate increase were observed (Figure 5).
The pH range in the overlying water of all cores and treatments was 6.5–7.9 in Oligo-R and 6.9–8.1 in Eutro-B, respectively (Figure 5). The pH of Eutro-B anoxic treatments gradually decreased from the start to the end of the incubation, with the anoxic, low temperature and anoxic, high temperature decreasing from 8 to 7.5 and 8 to 7, respectively. The redox potential in the overlying water was only measured in Eutro-B. The redox potential (converted to the standard hydrogen electrode potential) was in the range 386–406 mV for the oxic-low temperature treatment and 433–483 mV for the oxic-high temperature treatment. For the anoxic-low temperature treatment redox potential slightly decreased from 403 to 392 mV, whereas for the anoxic-high temperature treatment it decreased from 373 to 144 mV.
3.3. Solutes Fluxes
Solute fluxes in Eutro-B were markedly higher than in Oligo-R except for DOC and Mn which appear to be more reactive in sediments from Oligo-R (Table 2, Figure 6). Fluxes of Fe and P in both reservoirs were very low for the oxic-low, oxic-high, and anoxic-low temperature treatments while they were high for the anoxic-high temperature treatment (Table 2, Figure 6). For the anoxic-high temperature conditions, fluxes of SRP and DP in Eutro-B, were approximately 10 times higher than in Oligo-R, while those of Fe were approximately five times higher than in Oligo-R. Synergistic effects (i.e., a significant interaction between temperature and oxygen conditions) were detected for SRP and Fe in Eutro-B (DP close to significance) but could not be identified in Oligo-R (Table 3). The fluxes of Mn and DOC showed a similar pattern with highest flux rates in the anoxic-high temperature treatment but effective rates were higher in Oligo-R.
Nitrogen components (nitrate and ammonia) showed markedly higher fluxes in Eutro-B compared to Oligo-R and the responsiveness to oxygen conditions and temperature was complex. While in Eutro-B both N ANOVA terms were significant, only NH4+-N showed a significant interaction term in Oligo-R. Sulphate and nitrate concentrations decreased during the experiment, indicating nitrate and sulphate reduction. Sulphate fluxes in Eutro-B decreased independent from oxic status and temperature constantly by about 20% from approx. 60 mg L−1 to 48 mg L−1. In contrast, a sulphate decrease in Oligo-R only occurred under anoxic conditions. Ammonium release was also prominent under anoxic-high temperature conditions; however, this was only in Eutro-B (Table 2, Figure 6). Oxygen consumption was only measured in Bautzen in the oxic treatments and doubled when temperature increased from 6 °C to 20 °C.
In Eutro-B, DOC, SRP, DP, and NO3−-N components reacted more strongly to a change in temperature compared to a change in oxygen conditions, as indicated by partial R2 values (Table 3). The opposite was observed for Mn and NH4+-N showed a comparable magnitude of reactivity between temperature and oxygen conditions. In Oligo-R, SRP, DP, and SO42− were influenced more by a change in temperature as compared to a change in oxygen conditions, as indicated by partial R2 values (Table 3). Similarly to Eutro-B, the opposite was observed for the metals (Fe and Mn) that appear to be influenced mostly by oxygen conditions while DOC was under equal control from temperature and oxygen conditions.
3.4. Phosphorus Fractionation
The TP concentration in the upper 10 cm of both reservoirs was generally constant but TP content in Eutro-B was twice that in Oligo-R (Figure 6). Sediment P (upper 1 cm) in Eutro-B was dominated by redox sensitive BD-P which constitutes 52% of the P species (Figure 7). The second most abundant fractions constitute organic bound NaOH-NRP (35%) while all other fractions remained low: NH4Cl-P (4%), HCl-P (2%) and refractory residual-P (7%). Also the upper 1 cm sediment in Rappbode was dominated by organic bound NaOH-NRP (62.5%) and redox sensitive BD-P (31.2%), but here redox-bound P was not the dominant fraction. All other species remained, again, low: NH4Cl-P (0.2%), HCl-P (2%) and refractory residual P (4.1%). Metal oxide bound NaOH-SRP was negligible in both reservoirs. Both Eutro-B and Oligo-R showed an increasing organic bound NaOH-NRP and decreasing redox sensitive BD-P with depth.
4. Discussion
4.1. Synergistic Temperature and Oxygen Condition Interaction
The highest P mobilization occurred under anoxic-high temperature conditions, which confirms our hypothesis that warming and anoxia synergistically intensify benthic release rate of different solutes. Higher temperatures increase the reduction of electron acceptors hence promotes a faster decrease in redox potential. In the Bautzen Reservoir, for example, redox potential in the anoxic-low temperature (392 mV) was higher than in the anoxic-high temperature (142 mV) cores, at the end of the experiment. Mobilization of solutes (P, NH4+-N, DOC) under oxic conditions was probably from aerobic degradation of organic matter [2] as revealed by the high amount of NaOH-NRP in the upper sediment layer (Figure 7). Under anoxia, two processes of solute mobilization were taking place, anaerobic degradation of organic matter (e.g., by denitrification or sulphate reduction) and desorption from redox-dependent minerals. These two processes co-occurred and were very different to disentangle in experiments and under in situ conditions.
Temperature and oxygen conditions effects at high temperature are difficult to disentangle since redox potential is also driven by temperature [1]. Pure oxygen conditions effects could only be realized by comparing the oxic-low and anoxic-low temperature treatments. Surprisingly, P release under anoxic-low temperature conditions was not different from P release under oxic-low temperature conditions. This was probably due to the presence of nitrate as shown by the relatively high redox potential in the anoxic-low temperature treatment. Sediment in Bautzen was collected in spring, therefore this sediment would require a far longer incubation to deplete nitrate. Nitrate prevents the reductive dissolution of Fe minerals bound to P [11,44] and hence, acts as a redox buffer as soon as oxygen is depleted. Our results might be an underestimation of the P release under long-term anoxic-low temperature conditions. Nevertheless, we can still draw an important conclusion; the presence of nitrate is critical in keeping the sediment oxidized after the depletion of oxygen and nitrate can buffer redox conditions over a long time as long as temperature is low. These interacting temperature–oxygen effects are particularly strong under high temperature when kinetics are fast, it is therefore interesting to analyze the conditions under low temperature in greater detail.
The pure temperature effect (constant oxygen conditions, different temperature) in our experiments can be derived from the solute flux differences between the oxic-low and oxic-high temperature treatments. Phosphorus mobilization was twice as higher under oxic-high than under oxic-low temperature conditions. This is interpreted as P mobilization from the temperature-dependent organic matter degradation [45,46]. However, P release from organic matter might be masked by the immobilization of released P by adsorption to Fe minerals as shown by the negative P flux in Oligo-R oxic-low temperature treatment. In such a case, the real mineralization flux of phosphorus would be even higher than that measured in our experimental setting.
The effects of temperature, oxygen and the interaction of temperature and oxygen on benthic solutes release were more visible for the eutrophic reservoir (Eutro-B) in comparison to the oligotrophic reservoir (Oligo-R, Table 3), obviously due to the lower trophic state and solute concentrations in Oligo-R. If we only consider Eutro-B (Table 3) we can see that the synergistic effects of temperature and oxygen conditions are prominent for N, P and, Fe and not for DOC and SO42− which is an indication of the high potential of nutrients release under warming of lakes.
4.2. Magnitude of Solutes Fluxes
The flux magnitude of NO3−-N, NH4+-N, SO42−, Fe, Mn and DOC was high under the anoxic high temperature conditions, with NH4+-N, DOC, Fe and Mn being released from the sediments, while NO3−-N and SO42− were reduced. Nitrate reduction was a third of the ammonium release indicating a supply of nitrogen from the sediments, most probably arising from mineralization of organic matter. Phosphorus and DOC are known to be associated with Fe [3,47] and their release confirms that P released from the sediment is to a large amount attributable to the iron-bound, redox dependent fraction. The observed sulphate reduction has implications on P-release by immobilization of Fe through formation of iron sulfides, which can intensify P release into the water and even mediate the persistence of P in the dissolved phase when the overlying water becomes oxic [48,49,50].
Phosphorus release patterns in the different treatment was similar in both the eutrophic and oligotrophic lake, however, the flux magnitude was different. The P fluxes in Eutro-B are 10 times higher than those in Oligo-R. Our direct measurements are explained by the higher percentage of BD-P in Eutro-B in comparison to Oligo-R. The BD-P fraction is the redox sensitive P which is bound to Fe and Mn compounds [40]. The high proportion of NaOH-NRP (62%) in Oligo-R is an indication of higher organic P. This implies a high potential P-release from mineralization processes in Oligo-R. However, this was not observed in our experiment and is most probably attributable to low organic matter quality and a high share of recalcitrant OM.
Fluxes of N just like those of P were generally higher in Eutro-B than Oligo-R. Nitrate reduction occurred in both reservoirs and in all treatments. The P fluxes from this study are within the same magnitude as from other sediment incubation studies (Table 4). Ammonium fluxes were positive in both reservoirs except for the anoxic-low temperature treatment in Oligo-R, where the NH4+-N flux was negative. Surprisingly positive NH4+-N fluxes were measured also under oxic conditions; normally, NH4+-N release is expected when overlying water is anoxic, since under oxic conditions NH4+-N released from the anoxic sediments will be oxidized to NO3−-N (nitrification). A sum of the NO3−-N and NH4+-N flux shows that there was a net positive N flux in both the oxic- and anoxic-high temperature treatments in Eutro-B which is an indication that warmer temperature will lead to mobilization of N from sediments even under oxic conditions. Our NH4+-N release rates are on the higher end of the scale in comparison to other studies (Table 4).
Sulphate reduction occurred under oxic conditions in Eutro-B, under anoxic conditions in Oligo-R, and a distinct decrease under anoxic conditions in Eutro-B. The observed sulphate decrease is most likely due to diffusion into the porewater. Although sulphate reduction under anoxic conditions is the most probable explanation for this decrease, adsorption of sulphate oxyanions on Fe(III) oxides/hydroxides of the sediment cannot be excluded, especially if reduced Fe(II) ions increase during the anoxic conditions [51]. Adsorption mechanisms of sulphate oxyanions onto Fe(III) oxides/hydroxides of the sediment are also presumed to be the reason for the slight decrease in sulphate concentrations under oxic conditions. Adsorption of sulphate oxyanions onto Fe(III) oxides/hydroxides is well studied since more than 30 years, e.g., [51,52,53]. Usually, adsorption of sulphate oxyanions onto Fe(III) oxides/hydroxides is low at circum-neutral pH but can be up to 25% if initial concentration was high (1 mM) [51]. The observed sulphate reduction under oxic conditions in Eutro-B might have led to immobilization of Fe and subsequent release of P as observed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 4.
Benthic phosphorus release in different lakes/reservoirs at different sediment incubation temperature and oxygen conditions.
Table 4.
Benthic phosphorus release in different lakes/reservoirs at different sediment incubation temperature and oxygen conditions.
| Lake | Trophic State | Incubation Conditions | Flux (mmol m−2 day−1) | Reference | ||
|---|---|---|---|---|---|---|
| Temp. (°C) | Oxygen Condition | SRP | NH4+-N | |||
| Lake Arreso, Denmark | Eutrophic | 18 °C | Oxic | 0.13–0.39 | ND | [54] |
| Spring Lake, USA | Eutrophic | 17–25 | Oxic | −0.07–0.01 | ND | [41] |
| 17–25 | Anoxic | 0.29–0.86 | ND | |||
| Grand Lake, USA | Oligotrophic | <16 °C | Oxic | −0.002 | ND | [55] |
| >16 °C | Oxic | 0.02 | ND | |||
| Chubb Lake, Canada | Oligotrophic | 11 °C | Anoxic | 0.07 | ND | [56] |
| Onodonga Lake, USA | Hyper-eutrophic | 4 °C | Oxic | 0.04 | ND | [1] |
| 4 °C | Anoxic | 0.39 | ND | |||
| 8 °C | Oxic | 0.20–0.21 | ND | |||
| 8 °C | Anoxic | 0.62–0.71 | ND | |||
| Lake Frances, USA | Hyper-eutrophic | 22 °C | Oxic | 0.12–0.31 | ND | [57] |
| 22 °C | Anoxic | 0.50–0.73 | ND | |||
| Loch Leven Lake, Scotland | Eutrophic | 17 °C | Oxic | 0.39 | ND | [58] |
| Acton Lake (reservoir) Ohio, USA | Eutrophic | 25 °C | Oxic | 0.03 | 0.95 | [59] |
| 12 °C | Hypoxic | 0.29 | 1.39 | |||
| Lake Arreskov | Eutrophic | 16 °C | Oxic | ~0.97 | ND | [11] |
| Lake Kvind | Eutrophic | 18 °C | Oxic | 1.58 | ND | |
| Lake Søbygård | Eutrophic | 20 °C | Oxic | 5.30 | ND | |
| Lake Væng | Eutrophic | <15 °C | Oxic | 1.13 | ND | |
| Lake Tahoe, California/Nevada, USA | Ultra-oligotrophic | 5 °C | Oxic | 0 | 0 | [60] |
| Anoxic | 0.009 | 0.04 | ||||
| Upper San Leandro Reservoir, Oakland | Meso/eutrophic | 12–16 °C | Anoxic | ND | 0.64 ± 0.4 | [61] |
| Lake Mathews, Riverside Reservoir | Oligo/mesotrophic | 12–16 °C | Anoxic | ND | 0.21 ± 0.07 | |
| Walker Lake | 12–16 °C | Anoxic | ND | 1.36 | ||
| Lafayette Reservoir, Lafayette | Hypereutrophic | 12–16 °C | Anoxic | ND | 1.29 ± 0.4 | |
| Lake Bard (Reservoir), Ventura | Mesotrophic | 12–16 °C | Anoxic | ND | 0.54 | |
| Upper Crystal Springs Reservoirs, San Francisco | Mesotrophic | 12–16 °C | Anoxic | ND | 0.11 ±0.05 | |
| Lower Crystal Springs Reservoirs, San Francisco | Oligo/mesotrophic | 12–16 °C | Anoxic | ND | 0.29 ± 0.05 | |
| San Andreas Reservoir, San Francisco | Oligo/mesotrophic | 12–16 °C | Anoxic | ND | 0.14 ± 0.04 | |
| San Antonio Reservoir, San Francisco East Bay | Meso/eutrophic | 12–16 °C | Anoxic | ND | 0.31 ± 0.05 | |
| Lake Elsinore, Riverside | Hypereutrophic | 12–16 °C | Anoxic | ND | 6.43 ± 1.93 | |
| San Vicente Reservoir, San Diego | Hypereutrophic | 12–16 °C | Anoxic | ND | 2.36 ± 1.71 | |
| Lake Groß-Glienicke | Eutrophic lake | 16 °C | Oxic | 1.22 | ND | [62] |
| Anoxic | 2.77 | 0.08 | ||||
| Lake Arendsee | Eutrophic | 10 °C | Oxic | 0.08 | [63] | |
| Bautzen reservoir | Eutrophic | 6 °C | Oxic | 0.02 ± 0.01 | 0.71 ± 0.8 | This study |
| 6 °C | Anoxic | 0.04 ± 0.02 | 0.99 ± 0.36 | |||
| 20 °C | Oxic | 0.01 ± 0.003 | 2.11 ± 2.24 | |||
| 20 °C | Anoxic | 0.16 ± 0.03 | 5.63 ± 1.50 | |||
| Rappbode reservoir | Oligotrophic | 6 °C | Oxic | −0.0007 ± 0.0009 | 1.11 ± 0.43 | |
| 6 °C | Anoxic | 0.001 ± 0.002 | −0.02 ± 0.08 | |||
| 20 °C | Oxic | 0.0021 ± 0.0025 | 0.32 ± 0.32 | |||
| 20 °C | Anoxic | 0.0128 ± 0.0095 | 0.94 ± 0.35 | |||
4.3. Implications
Temperature and oxygen are important factors affecting the sediment–water interactions for various solutes. Our results show that the responses in solute fluxes are site-specific and usually greater in eutrophic reservoirs than in oligotrophic ones. The eutrophic systems also show a greater sensitivity to changes in temperature and oxygen conditions than sediments from oligotrophic waters. However, aside of trophic state also local geology might plays a role. For example, iron plays a larger role in Bautzen reservoir while in Rappbode reservoir manganese and allochthonous OM are more important. Prior knowledge about trophic state and local geology can therefore provide valuable knowledge to predict the reactivity of the sediments in case of anoxia and/or warming. A larger survey in different lakes or reservoirs with contrasting geology and trophic state could therefore provide a good data basis for more generalizable statements. Nevertheless, despite our results being laboratory based and only focusing on two reservoirs, we hypothesize that a similar result would be found in other lakes/reservoirs in the temperate region. Increasing lake temperature will lead to increased internal loading as a result of the synergistic effects of temperature and redox potential leading to the mobilization of nutrients both in the organic pool and those bound to redox-dependent metals.
We provide evidence that temperature and oxygen conditions act synergistically on sediment– water interactions. This synergistic effect implies that with a rising temperature in the system, a change in oxygen conditions is becoming more influential on, e.g., nutrient release. Generally speaking, temperature accelerates the sediment–water interactions. If we assume that a certain pulse of phosphorus can push a given lake or reservoir in a higher trophic level, e.g., from oligotrophic to eutrophic, we can expect that a critical temperature can be defined where a change in oxygen conditions is provoking this shift. Whenever a water body is entering a phase of hypolimnetic warming [10], e.g., because of climate warming or specific reservoir operation strategies, an increasing risk of eutrophication is arising. This risk is becoming real as soon as oxygen conditions in the hypolimnion deteriorate. Empirical studies on Lake Zürich exemplified that these mechanisms are relevant and taking place [16]. The supply of nutrients from the sediment is therefore an ongoing prominent feature of lakes/reservoirs under global warming and quantitative studies on the resulting solute fluxes are important for long-term lake management and climate adaptation.
Author Contributions
All the authors: T.D., K.R. and K.F. contributed substantially to this manuscript. Conceptualization–K.F. and K.R., Field sampling and laboratory work—T.D. and K.F., Data analysis—T.D. and K.F., Statistics—K.R., Manuscript preparation—T.D. and K.F., Manuscript review and editing—T.D., K.R., and K.F. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors would like to thank Michael Herzog, Karsten Rahn, and Martin Wieprecht for assistance with field sampling. We also thank Sebastian Lohr, Sebastian Back for conducting sediment incubations for Bautzen and Rappbode. We thank Andrea Hoff, Heike Goreczka, Ina Siebert, Kerstin Lerche, and Dorothee Ohlwein for chemical analyses. We appreciate the helpful comments of three anonymous reviewers.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Penn, M.R.; Auer, M.T.; Doerr, S.M.; Driscoll, C.T.; Brooks, C.M.; Effler, S.W. Seasonality in phosphorus release rates from the sediments of a hypereutrophic lake under a matrix of pH and redox conditions. Can. J. Fish. Aquat. Sci. 2000, 57, 1033–1041. [Google Scholar] [CrossRef]
- Hupfer, M.; Lewandowski, J. Oxygen controls the phosphorus release from lake sediments—A long-lasting paradigm in limnology. Int. Rev. Hydrobiol. 2008, 93, 415–432. [Google Scholar] [CrossRef]
- Dadi, T.; Friese, K.; Wendt-Potthoff, K.; Koschorreck, M. Benthic dissolved organic carbon fluxes in a drinking water reservoir. Limnol. Oceanogr. 2016, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, D.M.; Imboden, D.M. The prediction of hypolimnetic oxygen profiles: A plea for a deductive approach. Can. J. Fish. Aquat. Sci. 1996, 53, 924–932. [Google Scholar] [CrossRef]
- Seitzinger, S.P. Denitrification in freshwater and coastal marine ecosystems: Ecological and geochemical significance. Limnol. Oceanogr. 1988, 33, 702–724. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Internal phosphorus loading in shallow Danish lakes. Hydrobiologia 1999, 408, 145–152. [Google Scholar] [CrossRef]
- Gudasz, C.; Bastviken, D.; Steger, K.; Premke, K.; Sobek, S.; Tranvik, L.J. Temperature-controlled organic carbon mineralization in lake sediments. Nature 2010, 466, 478–481. [Google Scholar] [CrossRef]
- Kerimoglu, O.; Rinke, K. Stratification dynamics in a shallow reservoir under different hydro-meteorological scenarios and operational strategies. Water Resour. Res. 2013, 49, 7518–7527. [Google Scholar] [CrossRef]
- Lepori, F.; Roberts, J.J. Past and future warming of a deep European lake (Lake Lugano): What are the climatic drivers? J. Great Lakes Res. 2015, 41, 973–981. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Jagsch, A.; George, G.D.; Anneville, O.; Jankowski, T.; Wahl, B.; Lenhart, B.; Blenckner, T.; Teubner, K. Twenty years of spatially coherent deepwater warming in lakes across Europe related to the North Atlantic Oscillation. Limnol. Oceanogr. 2006, 51, 2787–2793. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.S.; Andersen, F.O. Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of four shallow, eutrophic lakes. Limnol. Oceanogr. 1992, 37, 577–589. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.-A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Alsterberg, C.; Hulth, S.; Sundbäck, K. Response of a shallow-water sediment system to warming. Limnol. Oceanogr. 2011, 56, 2147–2160. [Google Scholar] [CrossRef]
- Boehrer, B.; Schultze, M. Stratification of lakes. Rev. Geophys. 2008, 46, RG2005. [Google Scholar] [CrossRef] [Green Version]
- North, R.P.; North, R.L.; Livingstone, D.M.; Köster, O.; Kipfer, R. Long-term changes in hypoxia and soluble reactive phosphorus in the hypolimnion of a large temperate lake: Consequences of a climate regime shift. Glob. Chang. Biol. 2014, 20, 811–823. [Google Scholar] [CrossRef]
- Jean-Philippe, J.; Fabien, A.; Benjamin, A.; Jean-Marcel, D.; Pierre, S.; Michel, M.; Marie-Elodie, P. Inherited hypoxia: A new challenge for reoligotrophicated lakes under global warming. Glob. Biogeochem. Cycles 2014, 28, 1413–1423. [Google Scholar] [CrossRef]
- Fang, X.; Stefan, H.G. Simulations of climate effects on water temperature, dissolved oxygen, and ice and snow covers in lakes of the contiguous U.S. under past and future climate scenarios. Limnol. Oceanogr. 2009, 54, 2359–2370. [Google Scholar] [CrossRef]
- Rolighed, J.; Jeppesen, E.; Sondergaard, M.; Bjerring, R.; Janse, J.H.; Mooij, W.M.; Trolle, D. Climate Change Will Make Recovery from Eutrophication More Difficult in Shallow Danish Lake Sobygaard. Water 2016, 8, 459. [Google Scholar] [CrossRef] [Green Version]
- Einsele, W. Ueber die Beziehungen des Eisenkreislaufs zum Phosphatekreislauf im eutrophen See. Arch. Hydrobiol. 1936, 29, 664–686. [Google Scholar]
- Mortimer, C.H. The exchange of dissolved substances between mud and water in lakes. J. Ecol. 1942, 30, 147–201. [Google Scholar] [CrossRef]
- Wendt-Potthoff, K.; Kloß, C.; Schultze, M.; Koschorreck, M. Anaerobic metabolism of two hydro-morphological similar pre-dams under contrasting nutrient loading (Rappbode Reservoir System, Germany). Int. Rev. Hydrobiol. 2014, 99, 350–362. [Google Scholar] [CrossRef]
- Kelderman, P. Sediment-water exchange in Lake Grevelingen under different environmental conditions. Neth. J. Sea Res. 1984, 18, 286–311. [Google Scholar] [CrossRef]
- Rozan, T.F.; Taillefert, M.; Trouwborst, R.E.; Glazer, B.T.; Ma, S.F.; Herszage, J.; Valdes, L.M.; Price, K.S.; Luther, G.W. Iron-sulfur-phosphorus cycling in the sediments of a shallow coastal bay: Implications for sediment nutrient release and benthic macroalgal blooms. Limnol. Oceanogr. 2002, 47, 1346–1354. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wen, Y.; Zhou, J.; Wu, Y. Phosphorus release from lake sediments: Effects of pH, temperature and dissolved oxygen. KSCE J. Civ. Eng. 2014, 18, 323–329. [Google Scholar] [CrossRef]
- Sobek, S.; Cristian, G.; Birgit, K.; Tranvik, L.J.; David, B.; María, M.-P. Temperature Dependence of Apparent Respiratory Quotients and Oxygen Penetration Depth in Contrasting Lake Sediments. J. Geophys. Res. Biogeosci. 2017, 122, 3076–3087. [Google Scholar] [CrossRef] [Green Version]
- Liikanen, A.; Murtoniemi, T.; Tanskanen, H.; Väisänen, T.; Martikainen, P.J. Effects of Temperature and Oxygen Availability on Greenhouse Gas and Nutrient Dynamics in Sediment of a Eutrophic Mid-Boreal Lake. Biogeochemistry 2002, 59, 269–286. [Google Scholar] [CrossRef]
- Wouters, R. Das Bodewerk. Wasser Abfall 2011, 9, 10–15. [Google Scholar] [CrossRef]
- Bocaniov, S.A.; Barton, D.R.; Schiff, S.L.; Smith, R.E.H. Impact of tributary DOM and nutrient inputs on the nearshore ecology of a large, oligotrophic lake (Georgian Bay, Lake Huron, Canada). Aquat. Sci. 2013, 75, 321–332. [Google Scholar] [CrossRef]
- Rinke, K.; Kuehn, B.; Bocaniov, S.; Wendt-Potthoff, K.; Buettner, O.; Tittel, J.; Schultze, M.; Herzsprung, P.; Roenicke, H.; Rink, K.; et al. Reservoirs as sentinels of catchments: The Rappbode Reservoir Observatory (Harz Mountains, Germany). Environ. Earth Sci. 2013, 69, 523–536. [Google Scholar] [CrossRef]
- Benndorf, J.; Kranich, J.; Mehner, T.; Wagner, A. Temperature impact on the midsummer decline of Daphnia galeata: An analysis of long-term data from the biomanipulated Bautzen Reservoir (Germany). Freshwat. Biol. 2001, 46, 199–211. [Google Scholar] [CrossRef]
- Hülsmann, S. Recruitment patterns of Daphnia: A key for understanding midsummer declines? Hydrobiologia 2003, 491, 35–46. [Google Scholar] [CrossRef]
- Rinke, K.; Hübner, I.; Petzoldt, T.; Rolinski, S.; König-Rinke, M.; Post, J.; Lorke, A.; Benndorf, J. How internal waves influence the vertical distribution of zooplankton. Freshwat. Biol. 2007, 52, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Al-Mukhtar, M.; Dunger, V.; Merkel, B. Runoff and sediment yield modeling by means of WEPP in the Bautzen dam catchment, Germany. Environ. Earth Sci. 2014, 72, 2051–2063. [Google Scholar] [CrossRef]
- Friese, K.; Schultze, M.; Boehrer, B.; Büttner, O.; Herzsprung, P.; Koschorreck, M.; Kuehn, B.; Rönicke, H.; Tittel, J.; Wendt-Potthoff, K.; et al. Ecological response of two hydro-morphological similar pre-dams to contrasting land-use in the Rappbode reservoir system (Germany). Int. Rev. Hydrobiol. 2014, 99, 335–349. [Google Scholar] [CrossRef]
- Wentzky, V.C.; Tittel, J.; Jäger, C.G.; Rinke, K. Mechanisms preventing a decrease in phytoplankton biomass after phosphorus reductions in a German drinking water reservoir—Results from more than 50 years of observation. Freshwat. Biol. 2018, 63, 1063–1076. [Google Scholar] [CrossRef]
- Dadi, T.; Völkner, C.; Koschorreck, M. A sediment core incubation method to measure the flux of dissolved organic carbon between sediment and water. J. Soils Sed. 2015, 15, 2350–2358. [Google Scholar] [CrossRef]
- Morgenstern, P.; Friese, K.; Wendt-Potthoff, K.; Wennrich, R. Bulk Chemistry Analysis of Sediments from Acid Mine Lakes by Means of Wavelength Dispersive X-ray Fluorescence. Mine Water Environ. 2001, 20, 105–113. [Google Scholar] [CrossRef]
- Psenner, R.; Pucsko, R.; Sager, M. Die Fraktionierung organischer und anorganischer Phosphorverbindungen von Sedimenten—Versuch einer Definition ökologisch wichtiger Fraktionen. [Fractionation of organic and inorganic phosphorus compounds in lake sediments: An attempt to characterize ecologically important fractions]. Arch. Hydrobiol. Suppl. 1984, 70, 111–155. [Google Scholar]
- Hupfer, M.; Gächter, R.; Giovanoli, R. Transformation of phosphorus species in settling seston and during early sediment diagenesis. Aquat. Sci. 1995, 57, 305–324. [Google Scholar] [CrossRef]
- Steinman, A.; Rediske, R.; Reddy, K.R. The reduction of internal phosphorus loading using alum in Spring Lake, Michigan. J. Environ. Qual. 2004, 33, 2040–2048. [Google Scholar] [CrossRef]
- R-Core-Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Groemping, U. Relative Importance for Linear Regression in R: The Package relaimpo. J. Stat. Softw. 2006, 17, 27. [Google Scholar] [CrossRef] [Green Version]
- Grüneberg, B.; Dadi, T.; Lindim, C.; Fischer, H. Effects of nitrogen and phosphorus load reduction on benthic phosphorus release in a riverine lake. Biogeochemistry 2014, 1–18. [Google Scholar] [CrossRef]
- Ahlgren, J.; Tranvik, L.; Gogoll, A.; Waldebäck, M.; Markides, K.; Rydin, E. Sediment Depth Attenuation of Biogenic Phosphorus Compounds Measured by 31P NMR. Environ. Sci. Technol. 2005, 39, 867–872. [Google Scholar] [CrossRef]
- Gudasz, C.; Sobek, S.; Bastviken, D.; Koehler, B.; Tranvik, L.J. Temperature sensitivity of organic carbon mineralization in contrasting lake sediments. J. Geophys. Res. Biogeosci. 2015, 120, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Skoog, A.C.; Arias-Esquivel, V.A. The effect of induced anoxia and reoxygenation on benthic fluxes of organic carbon, phosphate, iron, and manganese. Sci. Total Environ. 2009, 407, 6085–6092. [Google Scholar] [CrossRef]
- Caraco, N.F.; Cole, J.J.; Likens, G.E. Sulfate control of phosphorus availability in lakes. Hydrobiologia 1993, 253, 275–280. [Google Scholar] [CrossRef]
- Holmer, M.; Storkholm, P. Sulphate reduction and sulphur cycling in lake sediments: A review. Freshwat. Biol. 2001, 46, 431–451. [Google Scholar] [CrossRef]
- Rapin, A.; Grybos, M.; Rabiet, M.; Mourier, B.; Deluchat, V. Phosphorus mobility in dam reservoir affected by redox oscillations: An experimental study. J. Environ. Sci. 2018. [Google Scholar] [CrossRef]
- Hinkle, M.A.G.; Wang, Z.; Giammar, D.E.; Catalano, J.G. Interaction of Fe(II) with phosphate and sulfate on iron oxide surfaces. Geochim. Cosmochim. Acta 2015, 158, 130–146. [Google Scholar] [CrossRef]
- Parfitt, R.L.; Smart, R.S.C. The Mechanism of Sulfate Adsorption on Iron Oxides1. Soil Sci. Soc. Am. J. 1978, 42, 48–50. [Google Scholar] [CrossRef]
- Geelhoed, J.S.; Hiemstra, T.; Van Riemsdijk, W.H. Phosphate and sulfate adsorption on goethite: Single anion and competitive adsorption. Geochim. Cosmochim. Acta 1997, 61, 2389–2396. [Google Scholar] [CrossRef]
- Søndergaard, M.; Kristensen, P.; Jeppesen, E. Phosphorus release from resuspended sediment in the shallow and wind-exposed Lake Arresø, Denmark. Hydrobiologia 1992, 228, 91–99. [Google Scholar] [CrossRef]
- Anthony, J.L.; Lewis, W.M. Low boundary layer response and temperature dependence of nitrogen and phosphorus releases from oxic sediments of an oligotrophic lake. Aquat. Sci. 2012, 74, 611–617. [Google Scholar] [CrossRef]
- Nürnberg, G.K.; Shaw, M.; Dillon, P.J.; McQueen, D.J. Internal Phosphorus Load in an Oligotrophic Precambrian Shield Lake with an Anoxic Hypolimnion. Can. J. Fish. Aquat. Sci. 1986, 43, 574–580. [Google Scholar] [CrossRef]
- Haggard, B.E.; Soerens, T.S. Sediment phosphorus release at a small impoundment on the Illinois River, Arkansas and Oklahoma, USA. Ecol. Eng. 2006, 28, 280–287. [Google Scholar] [CrossRef]
- Spears, B.M.; Carvalho, L.; Perkins, R.; Kirika, A.; Paterson, D.M. Sediment phosphorus cycling in a large shallow lake: Spatio-temporal variation in phosphorus pools and release. Hydrobiologia 2007, 584, 37–48. [Google Scholar] [CrossRef]
- Nowlin, W.H.; Evarts, J.L.; Vanni, M.J. Release rates and potential fates of nitrogen and phosphorus from sediments in a eutrophic reservoir. Freshwat. Biol. 2005, 50, 301–322. [Google Scholar] [CrossRef]
- Beutel, M.W.; Horne, A.J. Nutrient Fluxes From Profundal Sediment of Ultra-Oligotrophic Lake Tahoe, California/Nevada: Implications for Water Quality and Management in a Changing Climate. Water Resour. Res. 2018, 54, 1549–1559. [Google Scholar] [CrossRef]
- Beutel, M.W. Inhibition of ammonia release from anoxic profundal sediments in lakes using hypolimnetic oxygenation. Ecol. Eng. 2006, 28, 271–279. [Google Scholar] [CrossRef]
- Kleeberg, A.; Herzog, C.; Hupfer, M. Redox sensitivity of iron in phosphorus binding does not impede lake restoration. Water Res. 2013, 47, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, J.; Laskov, C.; Hupfer, M. The relationship between Chironomus plumosus burrows and the spatial distribution of pore-water phosphate, iron and ammonium in lake sediments. Freshwat. Biol. 2007, 52, 331–343. [Google Scholar] [CrossRef]
Figure 1.
Water temperature and oxygen profiles in Rappbode (top) and Bautzen (bottom) in early spring (dotted line) and late summer (solid line).
Figure 1.
Water temperature and oxygen profiles in Rappbode (top) and Bautzen (bottom) in early spring (dotted line) and late summer (solid line).

Figure 2.
Water profiles prior to sediment sampling. Please note; the Rappbode profile shown here was taken at a point of 17 m depth, the maximum depth is 89 m.
Figure 2.
Water profiles prior to sediment sampling. Please note; the Rappbode profile shown here was taken at a point of 17 m depth, the maximum depth is 89 m.

Figure 3.
Overlying water (OW) and porewater solutes concentration.

Figure 4.
Sediment characteristics of the upper 10 cm layer. Please note that for total amount of Al, Mn and Fe determined by XRF-analysis the sediments were sectioned in 2 cm layers.
Figure 4.
Sediment characteristics of the upper 10 cm layer. Please note that for total amount of Al, Mn and Fe determined by XRF-analysis the sediments were sectioned in 2 cm layers.

Figure 5.
Oxygen and pH during the incubation.

Figure 6.
Nutrients (N and P) fluxes. Please note that all solute fluxes data are presented in Table 2.
Figure 6.
Nutrients (N and P) fluxes. Please note that all solute fluxes data are presented in Table 2.

Figure 7.
P-fractionation in the upper 10 cm sediment layer, Oligo-R (left) and Eutro-B (right).

Table 1.
Reservoirs morphometry, hydrological, catchment and water quality properties. Data compiled from various sources [8,28,30,31,32,33,34,35,36].
| Parameter | Oligo-R | Eutro-B |
|---|---|---|
| Volume (million m3) | 113 | 39 |
| Surface area (km2) | 3.95 | 5.33 |
| Catchment area (km2) | 116 | 310 |
| Catchment land use (%) | ||
| Forest | 75 | 16 |
| Grasslands | - | 38 |
| Agriculture | 20 | 34 |
| Urban settlements | 5 | 12 |
| Dam height (m) | 106 | 19.4 |
| Elevation (m a.s.l) | 423.6 | 167.5 |
| Mean depth (m) | 28.6 | 7.4 |
| Maximum depth (m) | 89 | 13.5 |
| Retention time (day) | 344 | 164 |
| TP (µg L−1) | <5 | 60–100 |
| TN (mg L−1) | <1 | 2 |
| Chlorophyll a (µg L−1) | <10 | 50–80 |
| Trophic state | Oligotrophic | Eutrophic |
Table 2.
Mean fluxes and standard deviation of all parameters. Please note: The fluxes for SRP and NH4+-N are the same data used in Figure 6. NA refers to not analyzed.
Table 2.
Mean fluxes and standard deviation of all parameters. Please note: The fluxes for SRP and NH4+-N are the same data used in Figure 6. NA refers to not analyzed.
| Parameter (mmol m−2 day−1) | Oligo-R | Eutro-B | ||||||
|---|---|---|---|---|---|---|---|---|
| Oxic 6 °C n = 3 | Oxic 20 °C n = 3 | Anoxic 6 °C n = 3 | Anoxic 20 °C n = 3 | Oxic 6 °C n = 3 | Oxic 20 °C n = 4 | Anoxic 6 °C n = 4 | Anoxic 20 °C n = 4 | |
| DP | −0.003 ± 0.004 | 0.008 ± 0.008 | 0.002 ± 0.004 | 0.045 ± 0.026 | 0.06 ± 0.03 | 0.14 ± 0.07 | 0.05 ± 0.01 | 0.59 ± 0.13 |
| SRP | −0.002 ± 0.102 | 0.006 ± 0.007 | 0.003 ±0.006 | 0.039 ± 0.029 | 0.06 ± 0.04 | 0.12 ± 0.05 | 0.04 ± 0.01 | 0.50 ± 0.09 |
| NH4+-N | 1.11 ± 0.43 | 0.32 ± 0.32 | −0.02 ± 0.08 | 0.94 ± 0.35 | 0.71 ± 0.8 | 2.11 ± 2.24 | 0.99 ± 0.36 | 5.63 ± 1.50 |
| NO3−-N | −0.53 ± 0.1 | −0.54 ± 0.3 | −0.51 ± 0.23 | −0.94 ± 0.16 | −1.54 ± 0.34 | −0.24 ± 0.97 | −2.47 ± 0.16 | −1.24 ± 0.39 |
| DOC | −0.75 ± 0.27 | 0.94 ± 0.93 | 1.26 ± 1.14 | 10.39 ± 6.68 | 0.10 ± 0.18 | 1.89 ± 0.78 | 0.29 ± 0.33 | 1.78 ± 0.49 |
| Fe | −0.01 ± 0.01 | −0.01 ± 0.01 | 0.03 ± 0.06 | 0.20 ± 0.12 | 0.01 ± 0.01 | 0.001 ± 0.002 | 0.03 ± 0.02 | 1.09 ± 0.54 |
| Mn | −0.29 ± 0.19 | 0.22 ± 0.56 | 1.4 ± 1.05 | 5.16 ± 1.01 | −0.002 ± 0.03 | −0.02 ± 0.01 | 0.17 ± 0.06 | 0.15 ± 0.04 |
| SO42− | 0.24 ± 0.10 | 0.034 ± 0.12 | −0.05 ± 0.09 | −1.15 ± 0.08 | −1.88 ± 1.48 | −3.14 ± 2.39 | −1.78 ± 1.08 | −4.28 ± 1.78 |
| O2 | NA | NA | NA | NA | −28.53 ± 4.28 | −69.24 ± 23.14 | NA | NA |
Table 3.
Results from the statistical analyses of flux rates using ANOVA. Significant p-values for the two main effects (temperature and oxygen condition) and their interaction are printed in bold. The overall coefficient of determination (R2) of the underlying linear model is given in the last column. Partial R2 values are given in parentheses for each main factor and the interaction. Note that all partial R2 values sum up to the overall R2.
Table 3.
Results from the statistical analyses of flux rates using ANOVA. Significant p-values for the two main effects (temperature and oxygen condition) and their interaction are printed in bold. The overall coefficient of determination (R2) of the underlying linear model is given in the last column. Partial R2 values are given in parentheses for each main factor and the interaction. Note that all partial R2 values sum up to the overall R2.
| Variable | Temperature p-Value (Partial R2) | Oxygen Condition p-Value (Partial R2) | Interaction p-Value (Partial R2) | Overall R2 |
|---|---|---|---|---|
| Bautzen Reservoir (n = 15) | ||||
| SRP | 0.003 (0.380) | 0.025 (0.167) | 0.003 (0.153) | 0.70 |
| DP | 0.004 (0.366) | 0.031 (0.164) | 0.050 (0.144) | 0.67 |
| Fe | 0.007 (0.241) | 0.005 (0.272) | 0.009 (0.230) | 0.74 |
| Mn | 0.476 (0.018) | <0.001 (0.777) | 0.887 (0.000) | 0.80 |
| DOC | <0.001 (0.720) | 0.841 (0.001) | 0.556 (0.009) | 0.73 |
| NO3−-N | <0.001 (0.528) | <0.001 (0.368) | 0.007 (0.052) | 0.95 |
| NH4+-N | 0.002 (0.243) | <0.001 (0.347) | 0.003 (0.231) | 0.82 |
| SO42− | 0.05 (0.287) | 0.405 (0.038) | 0.800 (0.004) | 0.33 |
| Rappbode Reservoir (n = 12) | ||||
| SRP | 0.100 (0.231) | 0.155 (0.166) | 0.347 (0.067) | 0.46 |
| DP | 0.039 (0.292) | 0.065 (0.219) | 0.177 (0.105) | 0.62 |
| Fe | 0.132 (0.138) | 0.030 (0.341) | 0.147 (0.127) | 0.61 |
| Mn | 0.003 (0.224) | <0.001 (0.539) | 0.014 (0.130) | 0.89 |
| DOC | 0.056 (0.238) | 0.045 (0.268) | 0.163 (0.113) | 0.62 |
| NO3−-N | 0.185 (0.151) | 0.232 (0.120) | 0.179 (0.155) | 0.43 |
| NH4+-N | 0.716 (0.006) | 0.285 (0.054) | 0.005 (0.613) | 0.67 |
| SO42− | <0.001 (0.350) | <0.001 (0.453) | <0.001 (0.165) | 0.97 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dadi, T.; Rinke, K.; Friese, K. Trajectories of Sediment-Water Interactions in Reservoirs as a Result of Temperature and Oxygen Conditions. Water 2020, 12, 1065. https://doi.org/10.3390/w12041065
AMA Style
Dadi T, Rinke K, Friese K. Trajectories of Sediment-Water Interactions in Reservoirs as a Result of Temperature and Oxygen Conditions. Water. 2020; 12(4):1065. https://doi.org/10.3390/w12041065
Chicago/Turabian StyleDadi, Tallent, Karsten Rinke, and Kurt Friese. 2020. "Trajectories of Sediment-Water Interactions in Reservoirs as a Result of Temperature and Oxygen Conditions" Water 12, no. 4: 1065. https://doi.org/10.3390/w12041065
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.