Long-Term Changes of Species Composition and Functional Traits of Epiphytic Diatoms in the Szigetköz Region (Hungary) of the Danube River

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
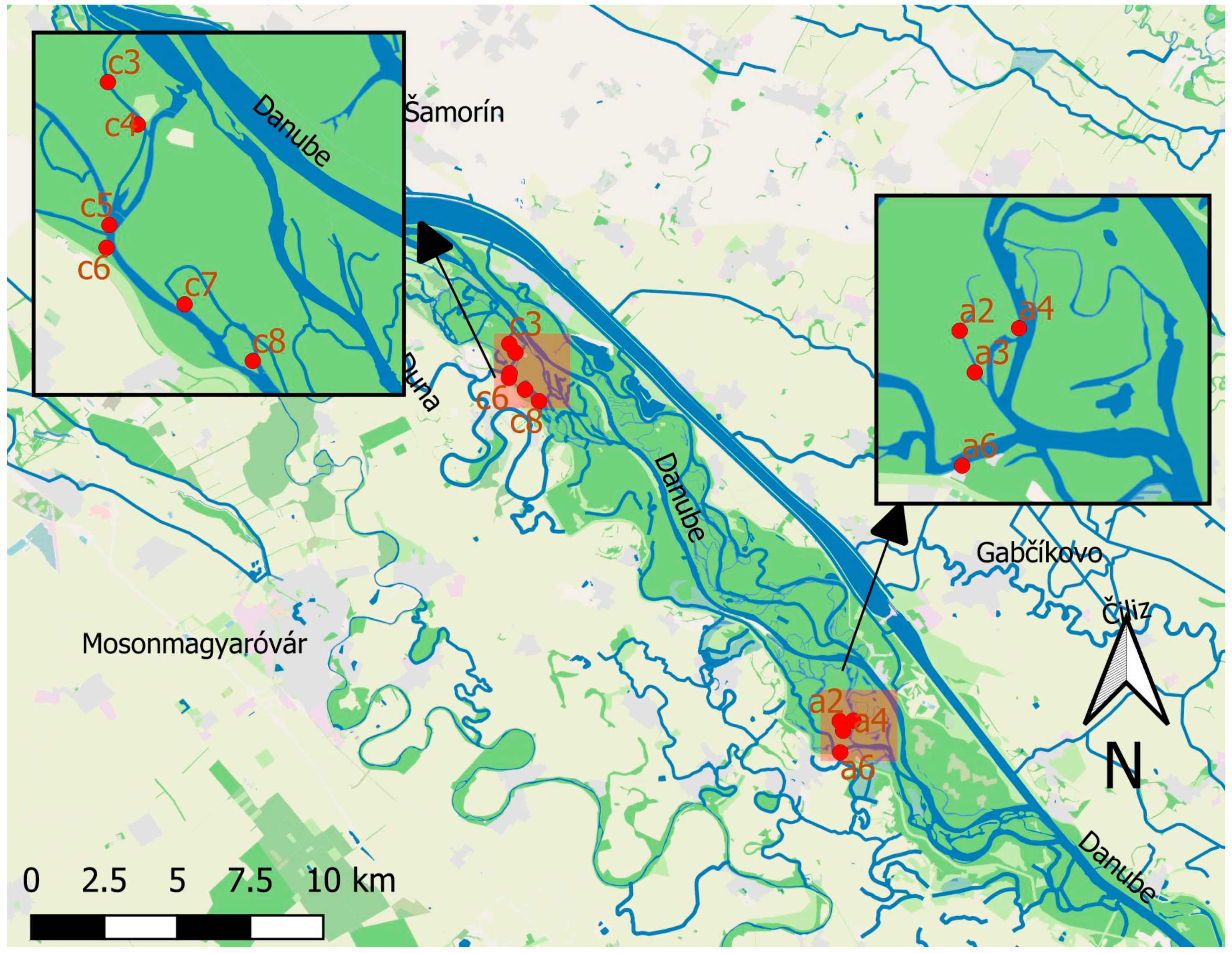
2.1. Study Site
2.2. Sampling Design
2.2.1. Sampling Design for Monitoring the Long-Term Changes
2.2.2. Sampling Design for Ecological Potential Assessment
2.3. Sample Processing
2.4. Diatom Index, Trait Data, Core and Common Species
2.5. Statistical Analyses
3. Results
3.1. Structure of the Diatom Assemblage
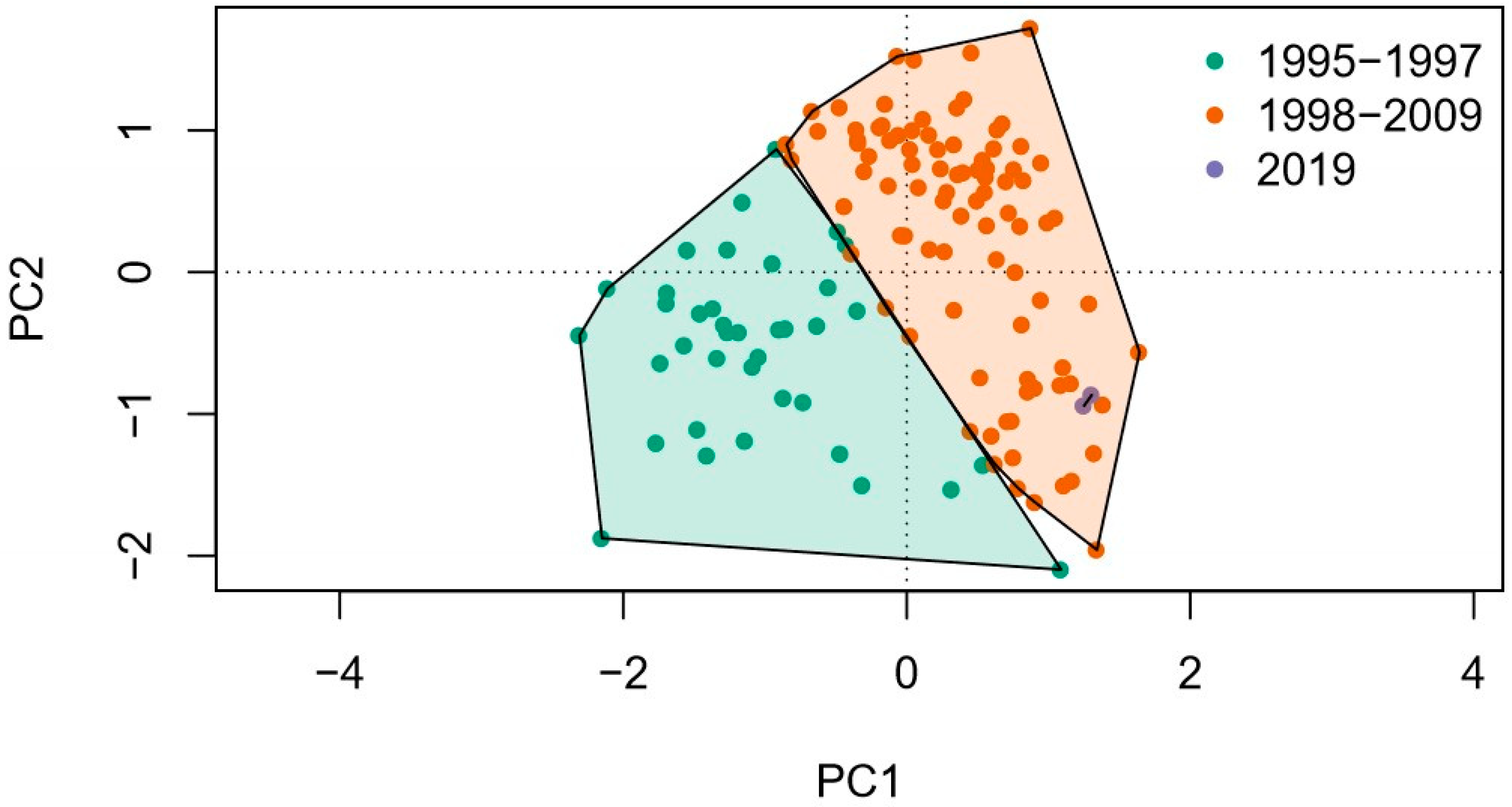
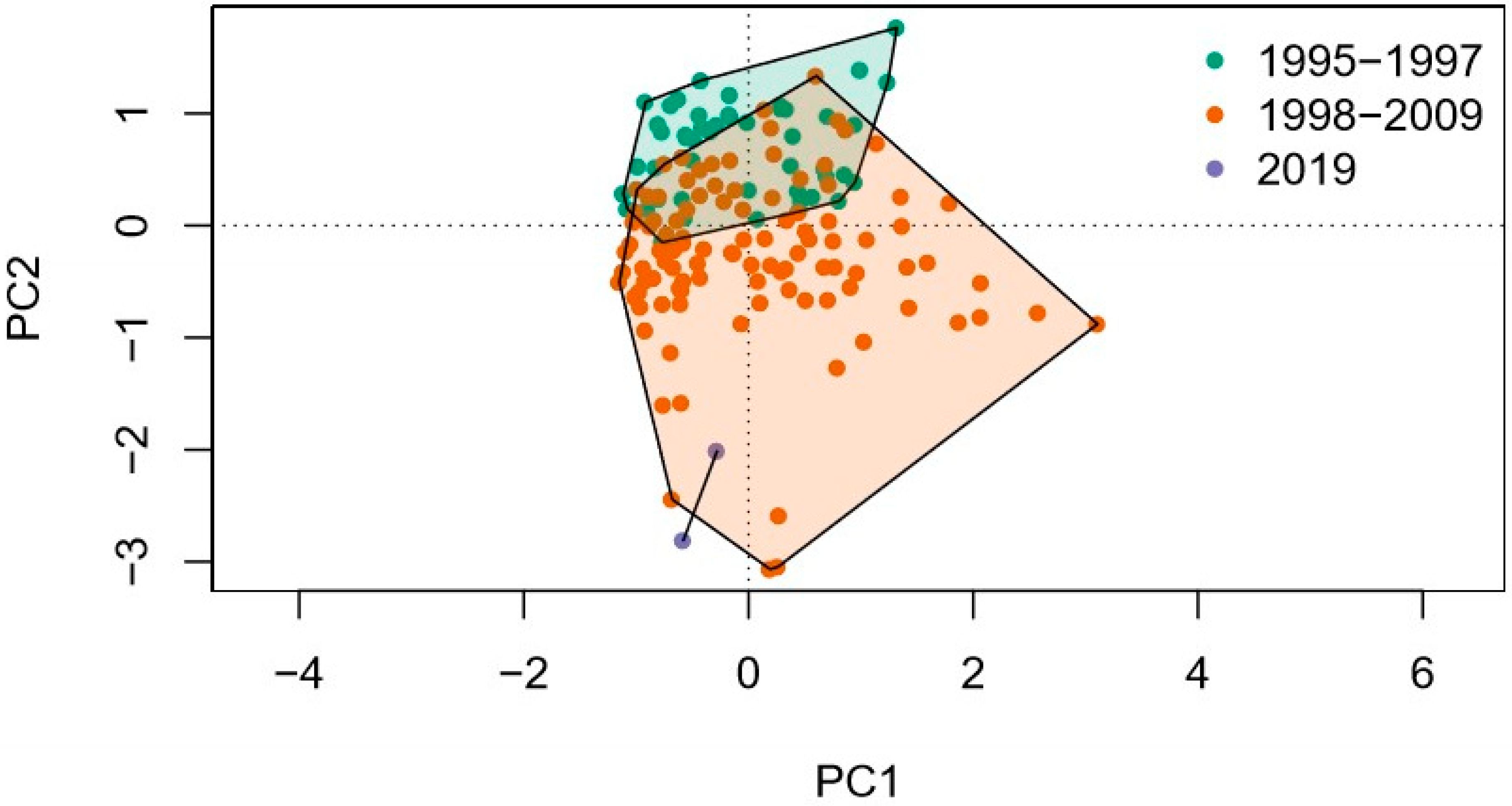
3.1.1. Differences of Diatom Assemblages before and after the Alteration of Water Quantity
3.1.2. Spatial Variance of the Diatom Assemblage Structure
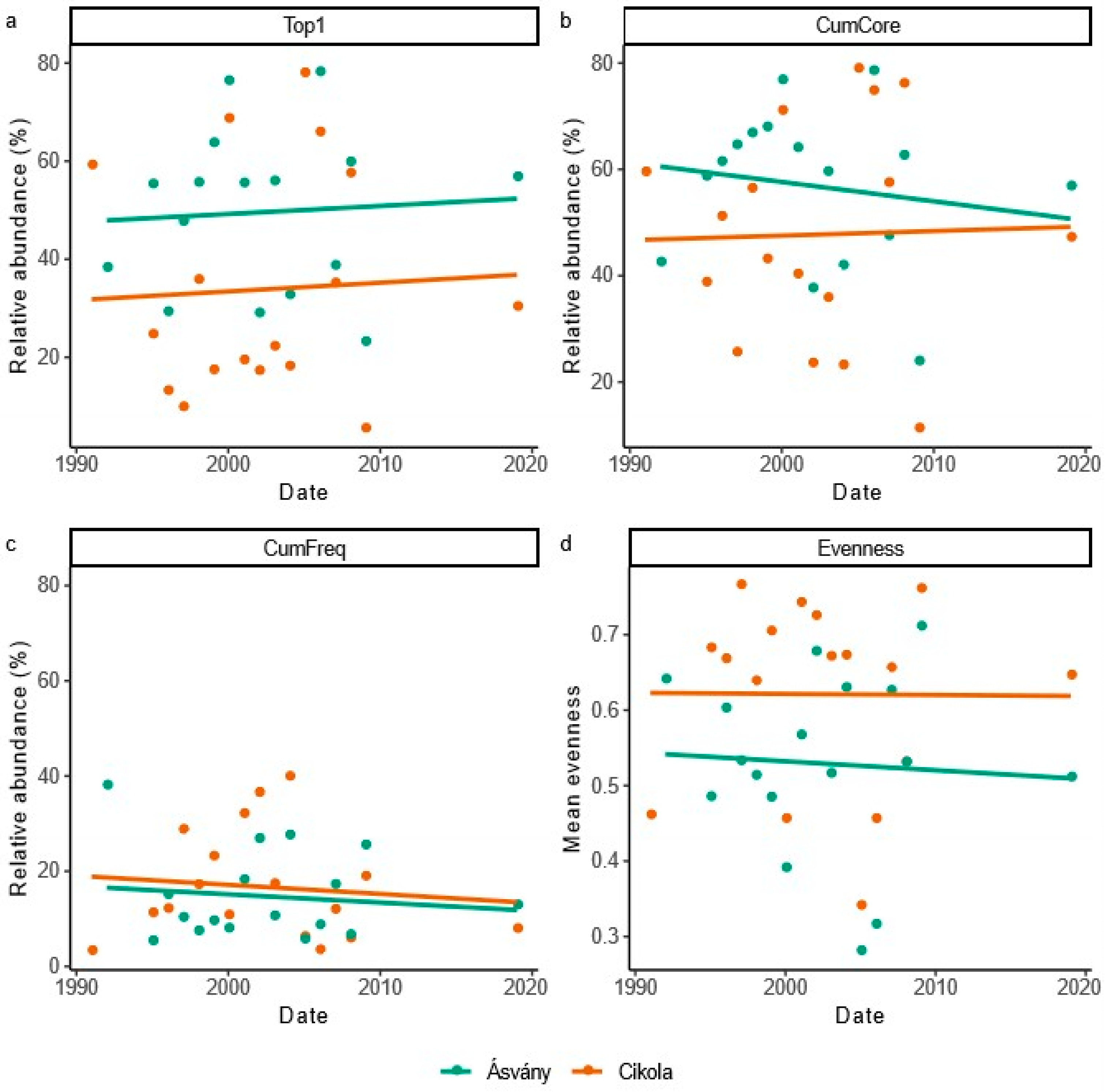
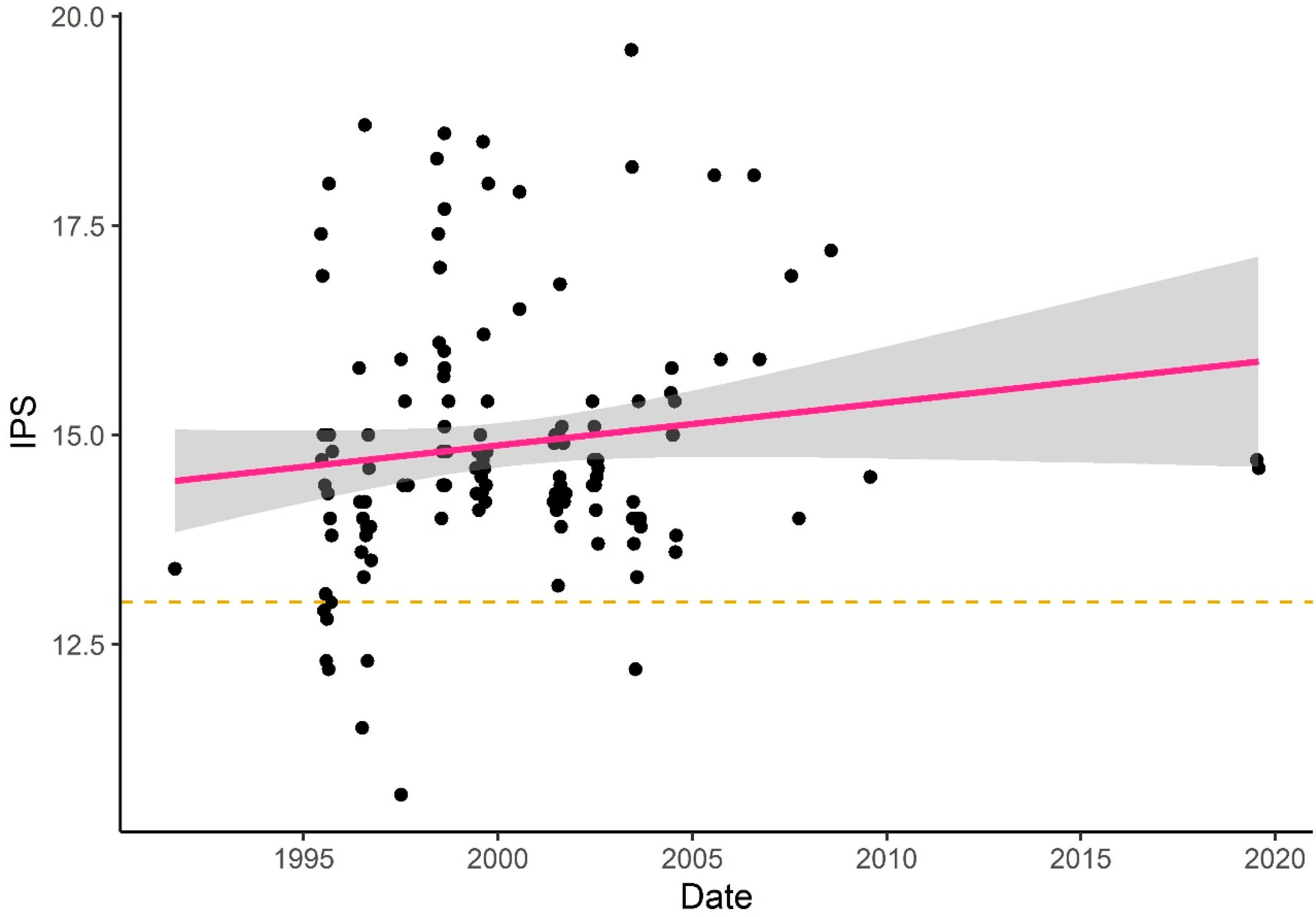
3.1.3. Temporal Variance of Diatom Assemblage Structure
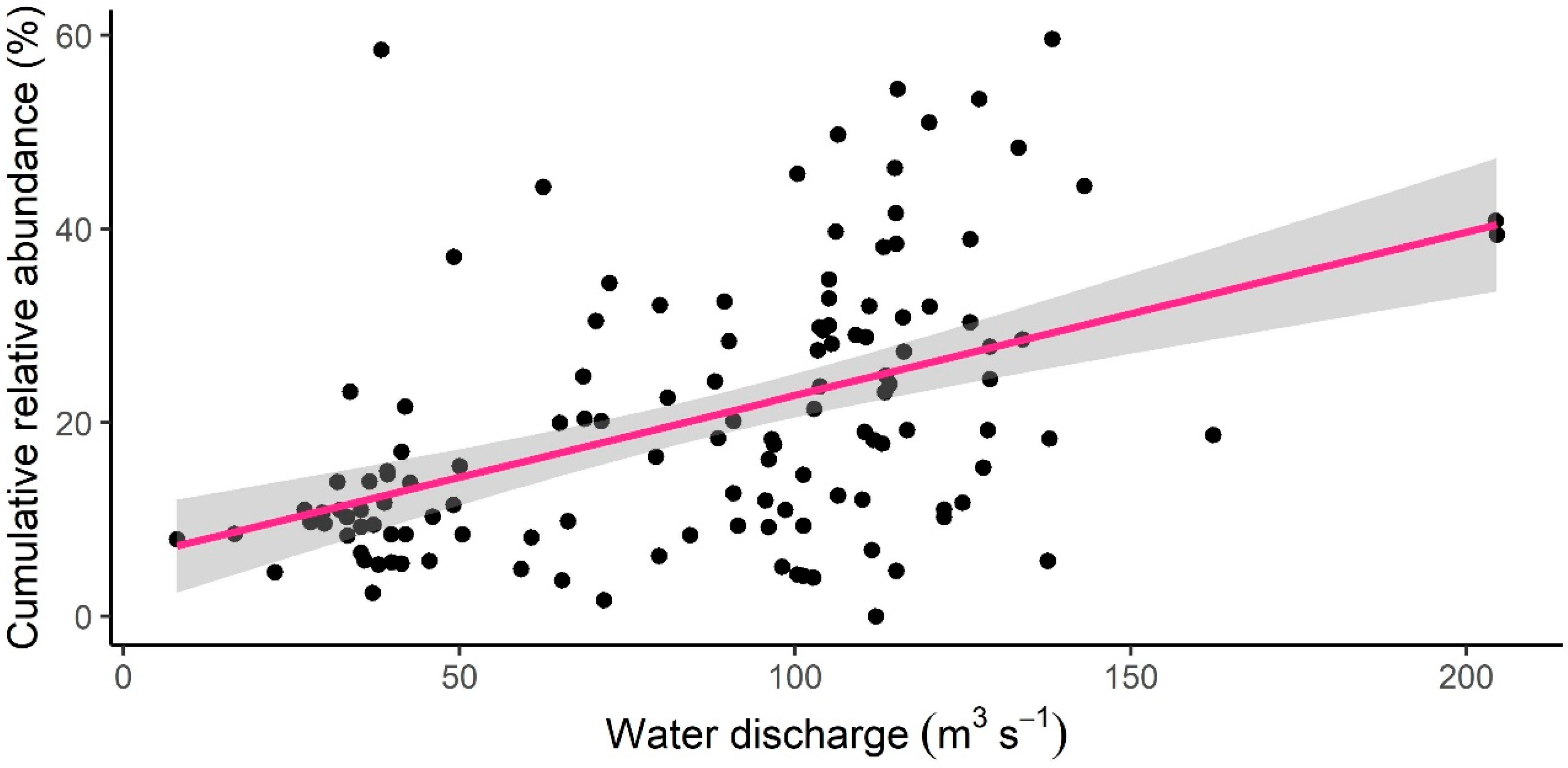
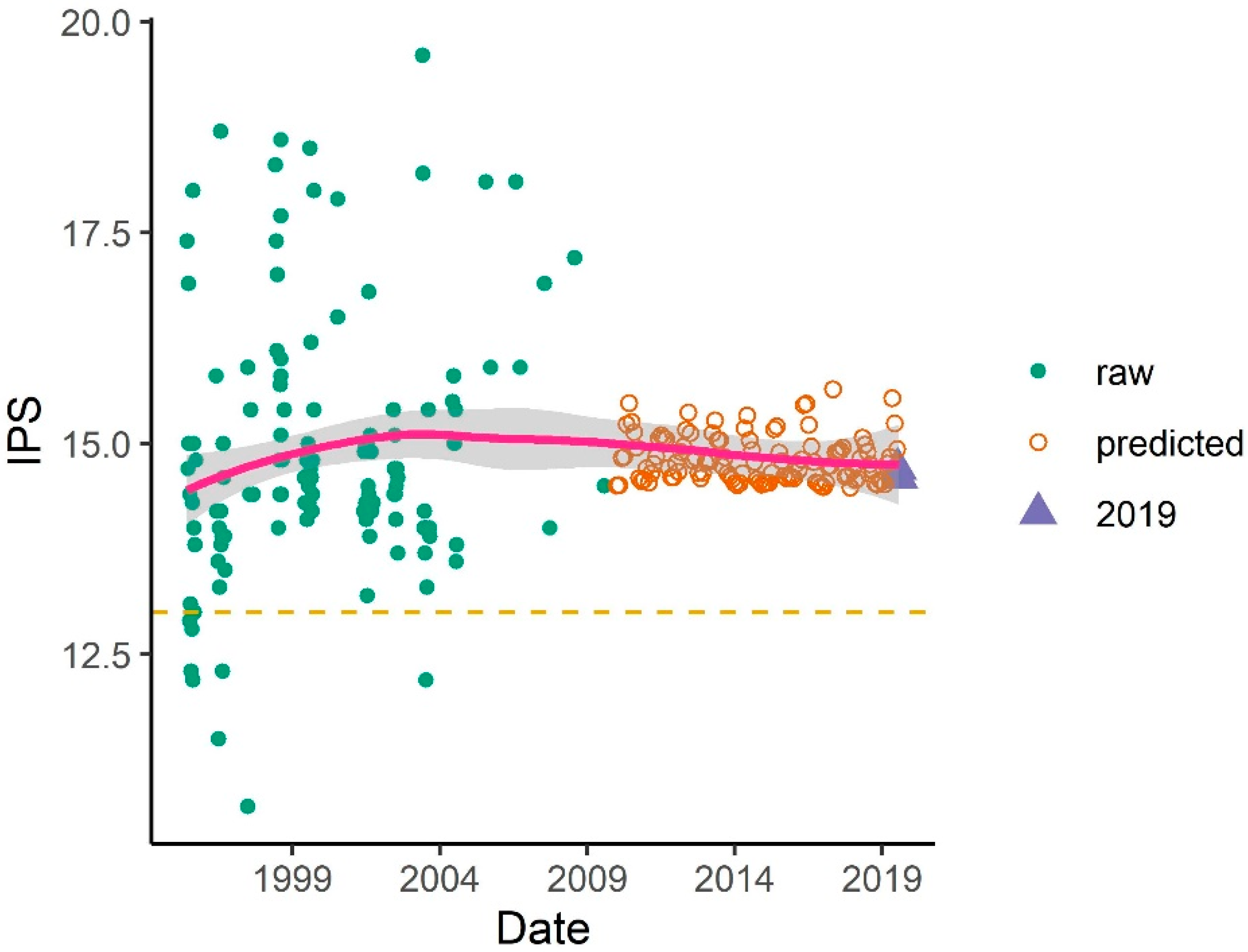
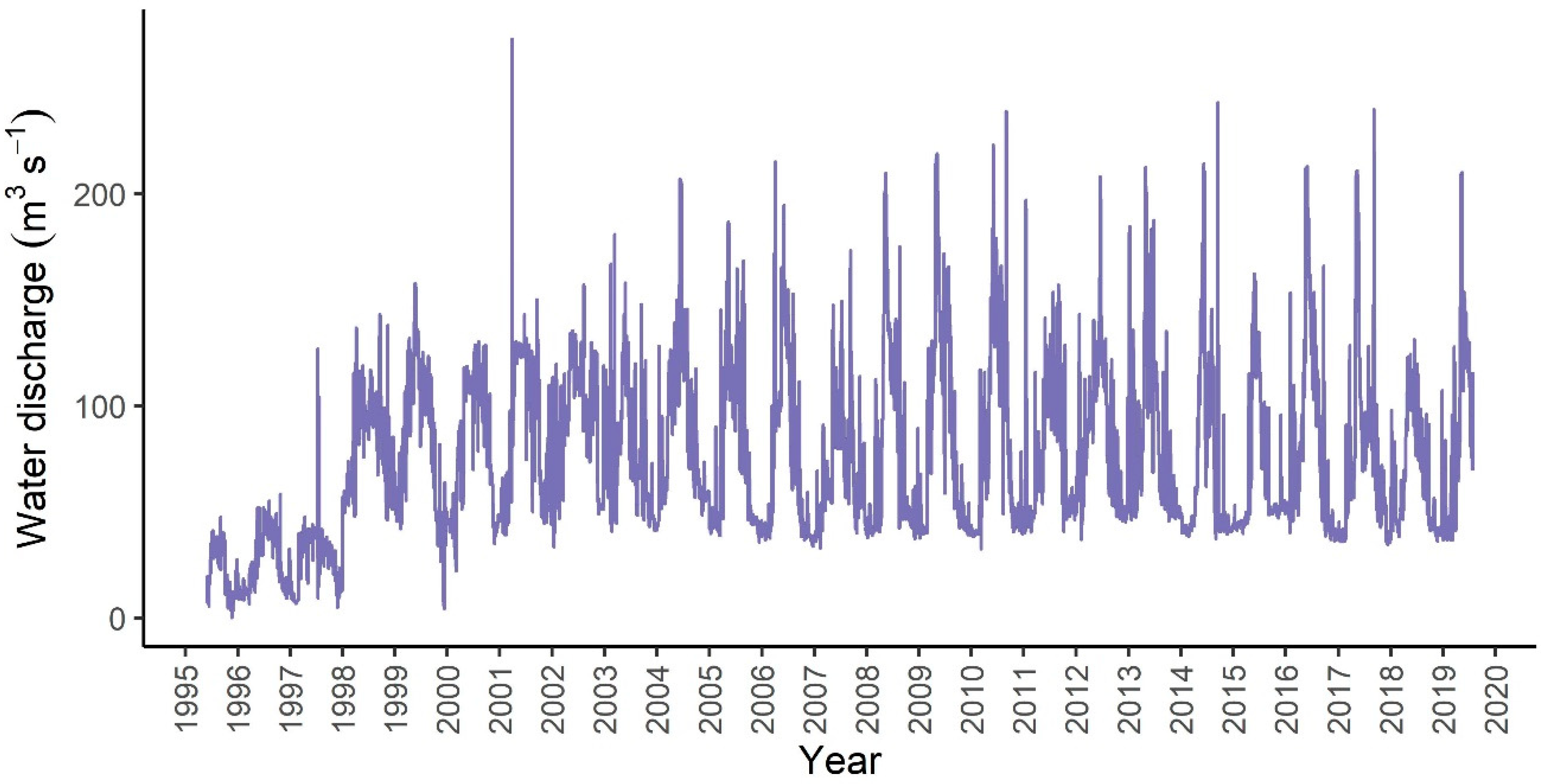
3.1.4. Correlation Between Water Discharge and Diatom Assemblage Structure
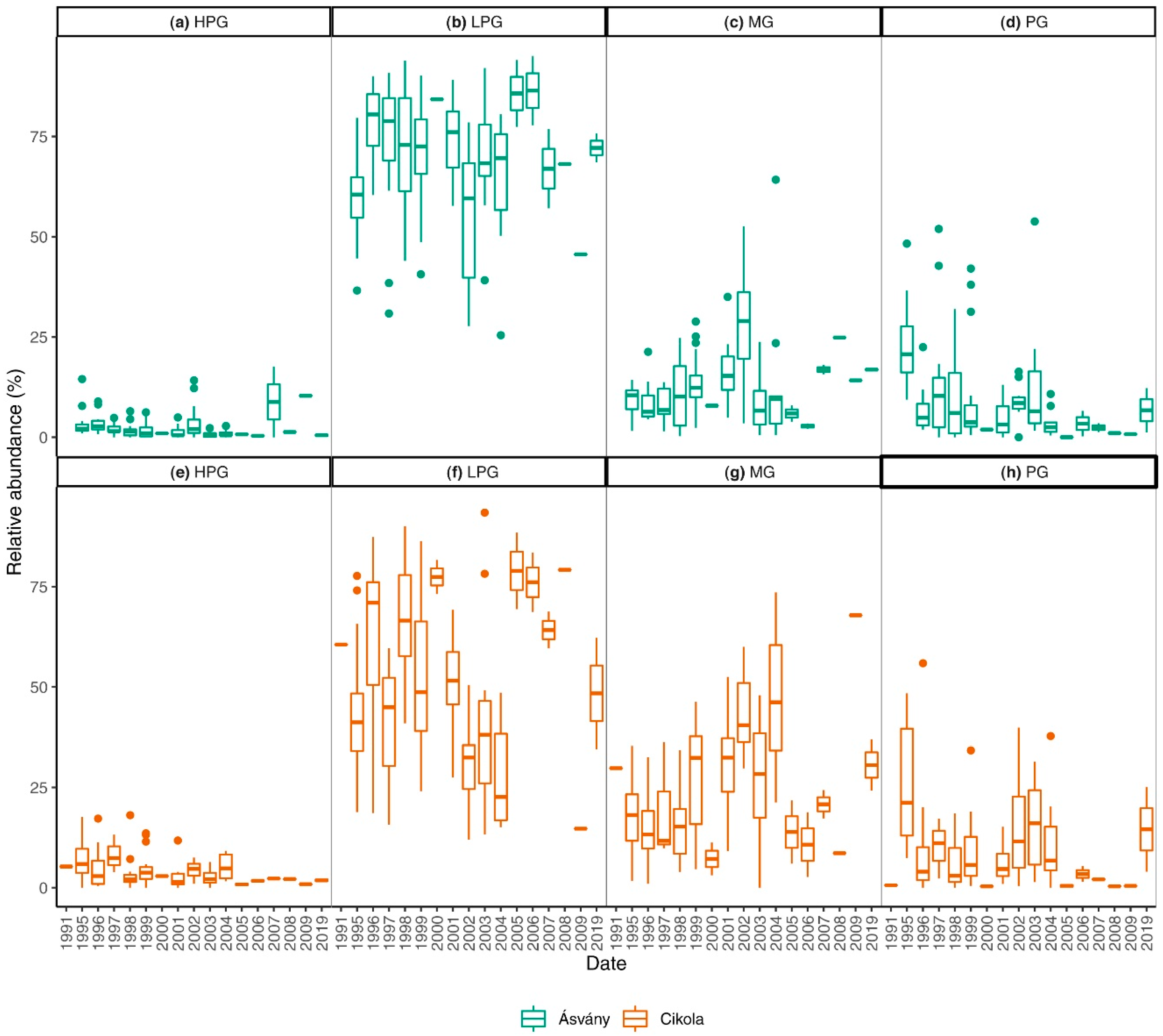
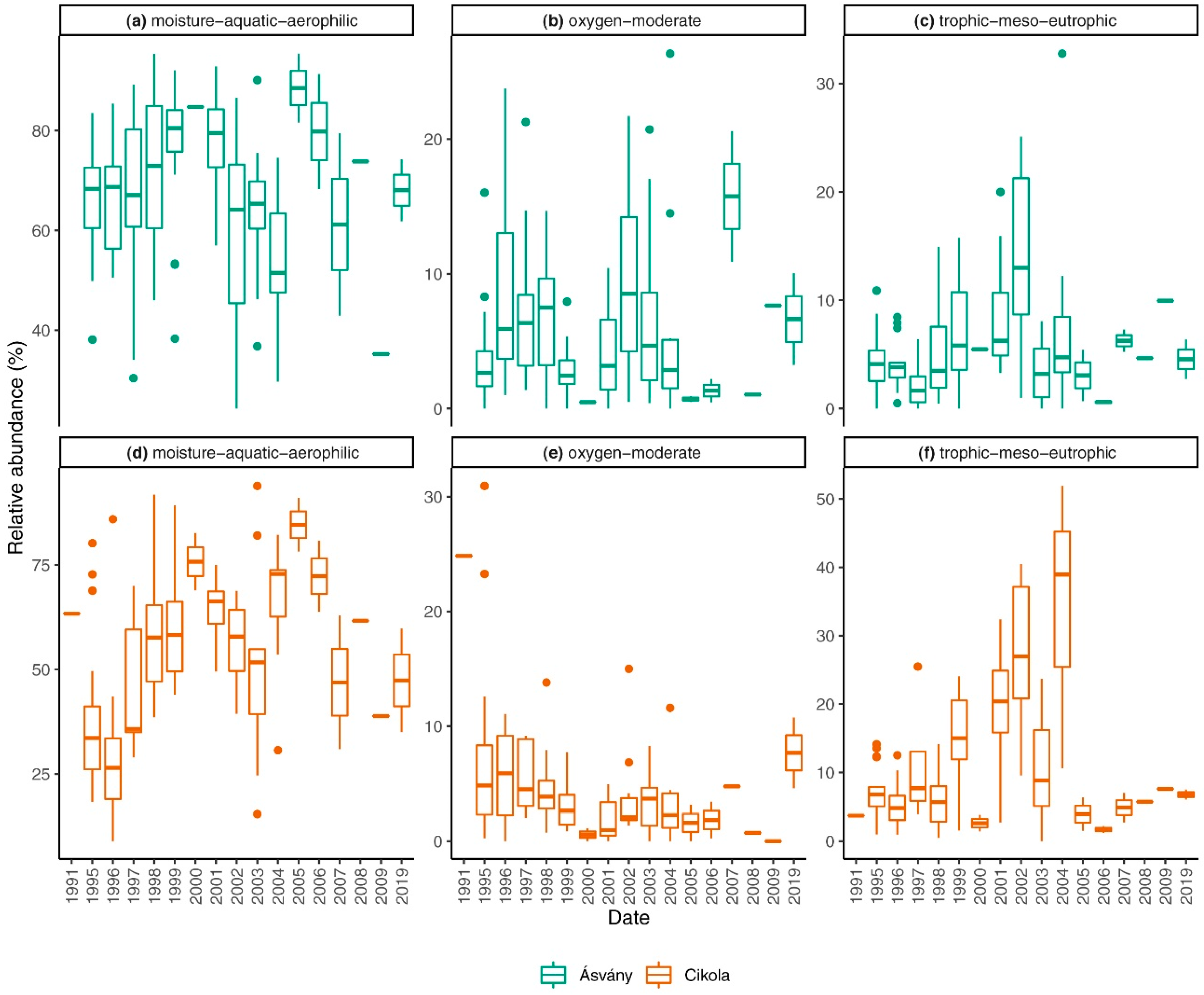
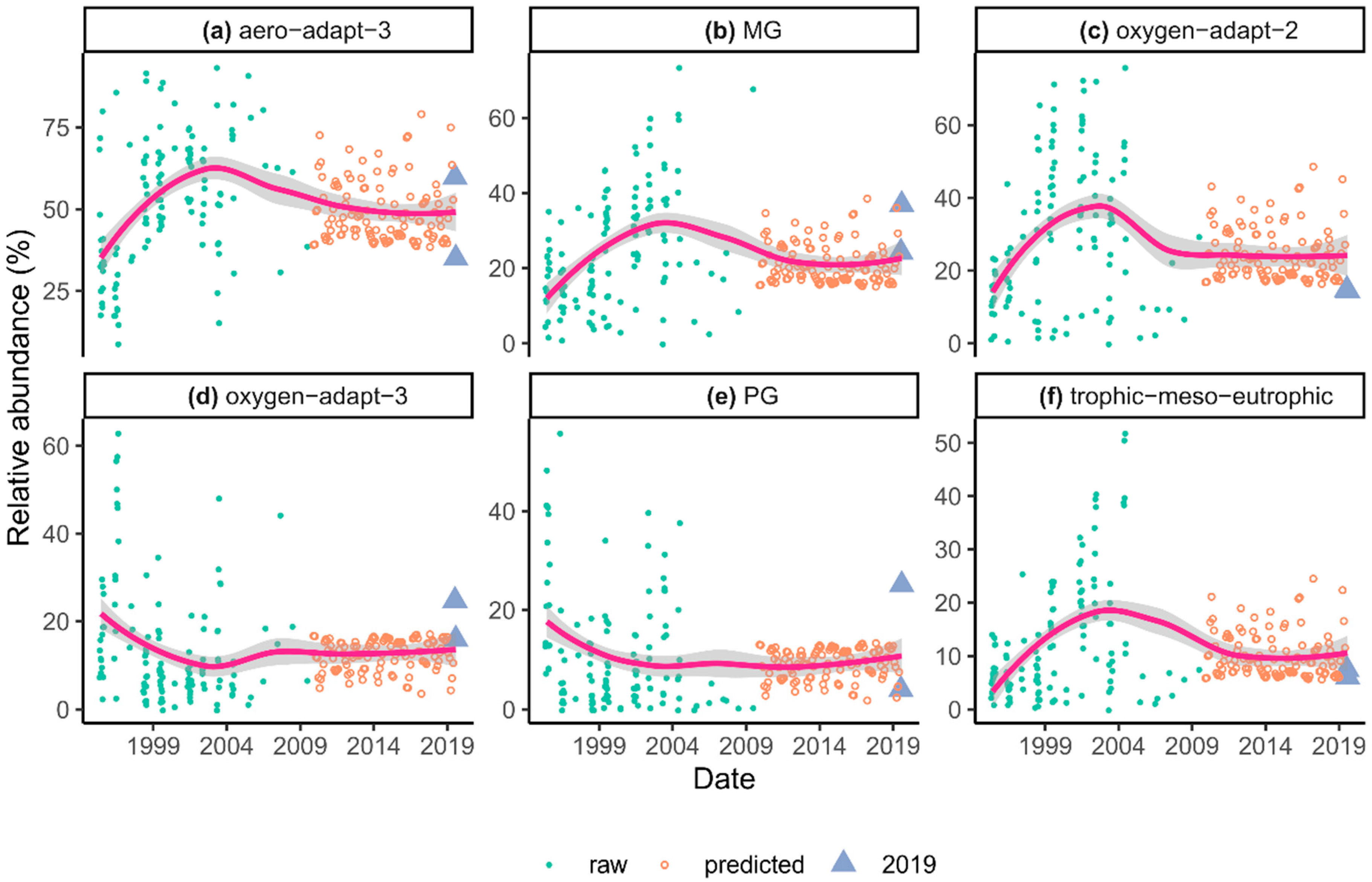
3.2. Traits of the Diatom Assemblage
3.3. Ecological Potential Assessment
3.3.1. Temporal Changes in the Eupotamic Branches of the Tributary Systems
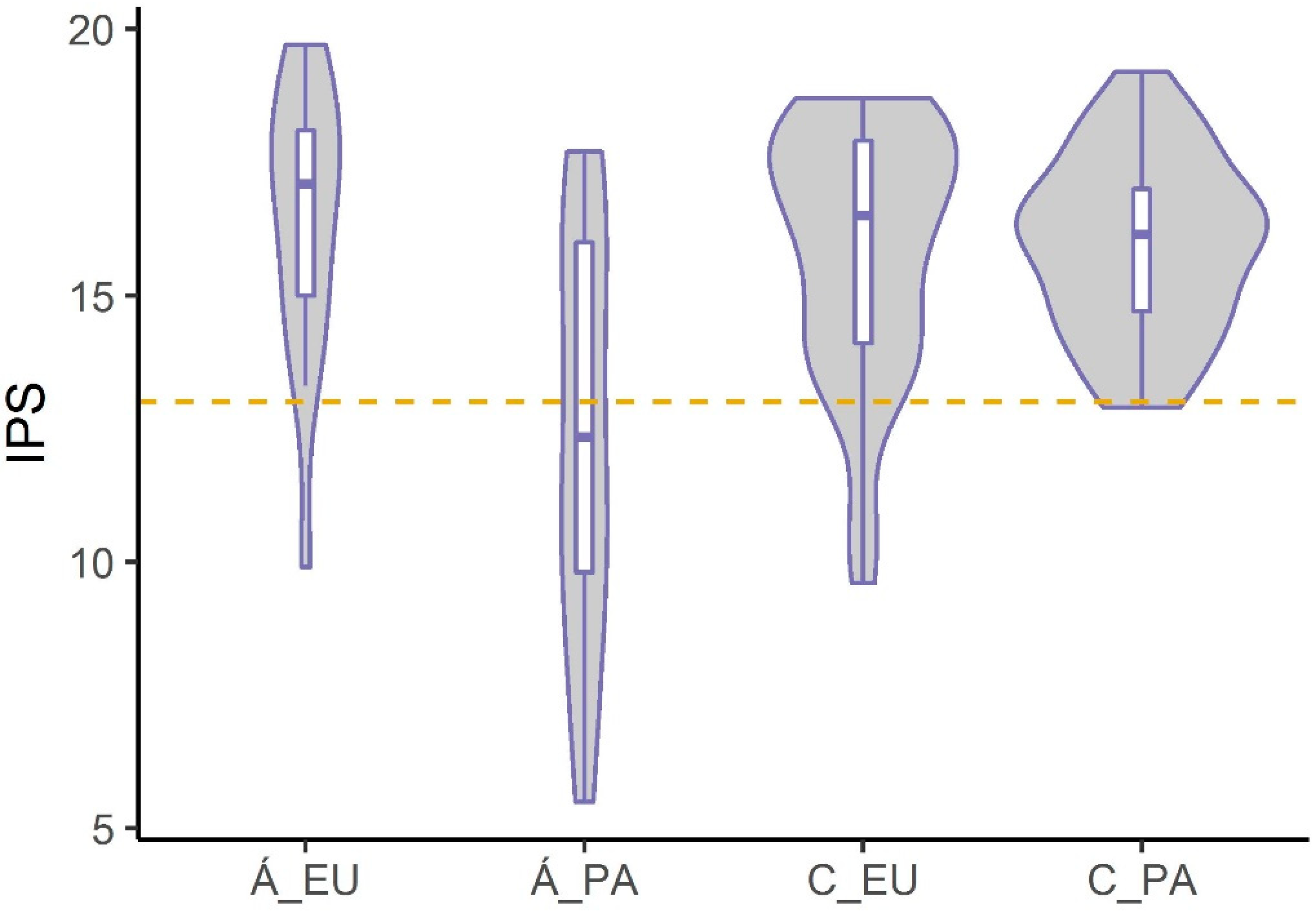
3.3.2. Differences in Eupotamic and Parapotamic Branches of the Tributary Systems
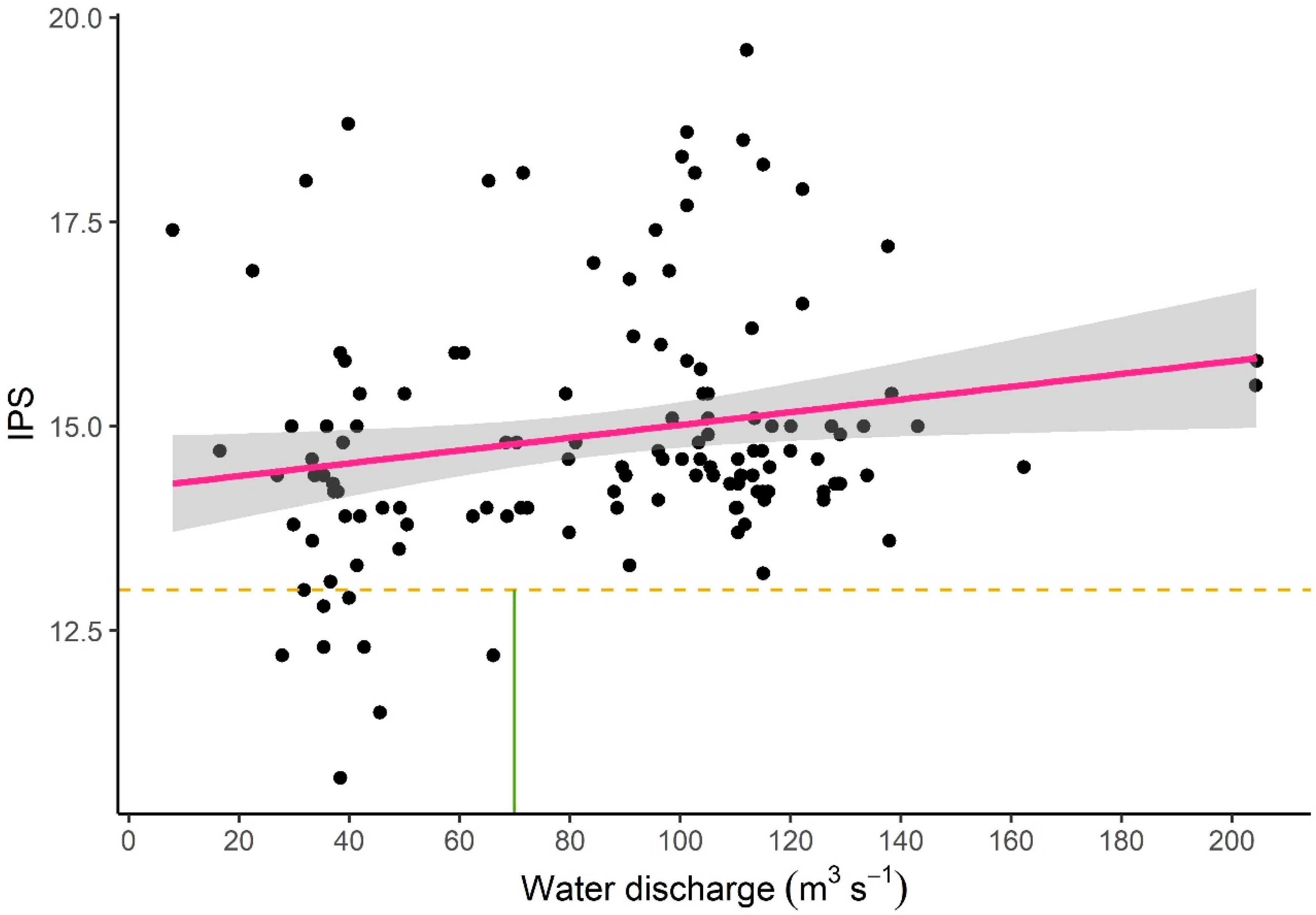
3.3.3. Correlation between Water Discharge and Ecological Potential
3.4. Model—Effect of Rehabilitation
4. Discussion
4.1. Structure of Diatom Assemblage
4.2. Traits of Diatom Assemblages
4.3. Ecological Potential Assessment
4.4. Model–Effect of Rehabilitation
5. Conclusions and Implications for Management
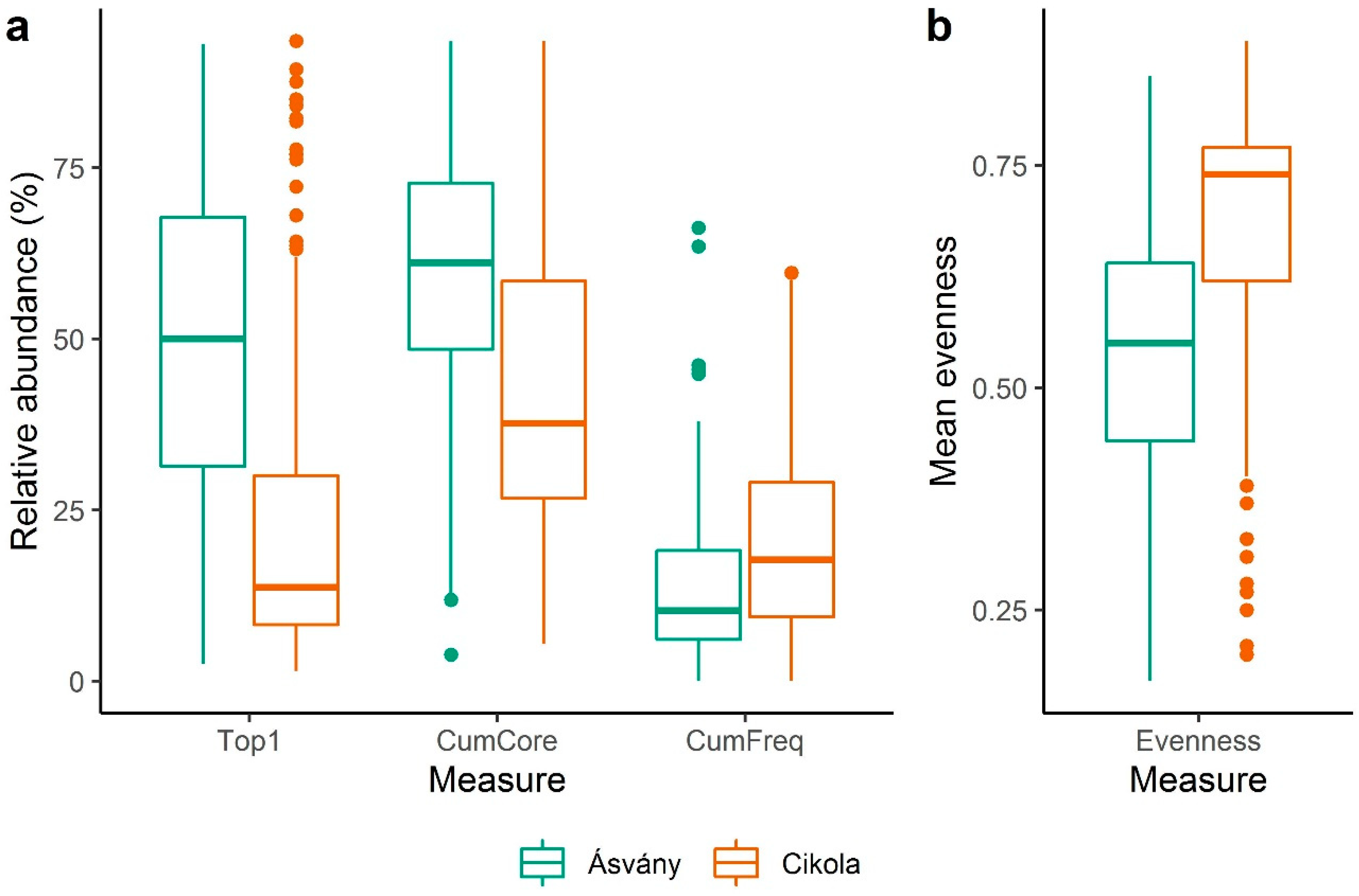
- Our results revealed greater spatial differences in the relative abundance of core species than in time due to spatial differences in environmental condition (first of all because of differences in water quantities between the Upper and Lower Szigetköz). The relative abundance of common species increased with increasing water discharge, as faster water flow could support species dispersion.
- After the diversion of Danube River, significant changes took place in diatom traits as well, but after the introduction of artificial water supply smaller changes occurred, especially for traits associated with water volume.
- Our results confirmed the need for rehabilitation of the Szigetköz tributary systems of the Danube, as rehabilitation improved the ecological potential of the branch system because of the increased water volume. There was a slight difference in the ecological potential of eupotamic and parapotamic branches in the rehabilitated Upper Szigetköz, while the ecological potential of parapotamic branches was lower than that of eupotamic ones in the Lower Szigetköz not rehabilitated at that time.
- In order to maintain good ecological potential, at least 70 m3 s−1 of water should be introduced into the branch system based on the phytobenthos study. Although, 70 m3 s−1 is necessary for good ecological potential, it is not per se a sufficient threshold. The method used here provides the basis for further targeted studies to clarify a lower threshold of good ecological potential and to fine-tune water management.
- Regular investigations and a research program are needed to better understand how communities respond to anthropogenic impacts and rehabilitation, as well as to predict how the ecosystem of the Szigetköz will respond to the effects of global warming and adapt to changing environmental conditions (temperature rise, extreme water regimes).
- Our long-term data sets may provide a good basis for comparing, for example, the abundance of core species and community structures in different aquatic ecosystems as they respond to different anthropogenic pressures (e.g., nutrient loads). Using such and similar data sets, we can understand and predict how algal core species will cope with the effects of accelerating climate change (temperature rise, altered water regime, water level fluctuation, etc.).
- Our results support the finding that different levels of organisms (both at species and trait levels) need to be explored to gain a deeper understanding of the role of environmental, climatic, regional, and temporal factors in the structure of biological communities [8].
- We studied a “natural mesocosm” site where different hydrological changes occurred in the same catchment area and these effects could be studied on traits. In light of the impact of global warming stressors on biodiversity, the identification of adaptive traits is of great importance.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Sites | |
|---|---|---|
| Top1 CV% mean | 48.1 | 74.7 |
| Cum. core CV% mean | 25.9 | 40.1 |
| Cum. com. CV% mean | 44.3 | 75.2 |
| Evenness CV% mean | 17.9 | 23.7 |
References
- Fryirs, K.A.; Brierley, G.J. Assessing the geomorphic recovery potential of rivers: Forecasting future trajectories of adjustment for use in management. WIREs Water 2016, 3, 727–748. [Google Scholar] [CrossRef]
- Ibáñez, C.; Peñuelas, J. Changing nutrients, changing rivers. Science 2019, 365, 637–638. [Google Scholar] [CrossRef] [PubMed]
- IPCC Intergovernmental Panel on Climate Change. Summary for policy makers, Technical summary. In AR5 Climate Change 2014: Mitigation of Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; Available online: https://www.ipcc.ch/report/ar5/wg3/ (accessed on 20 November 2019).
- Easterling, D.R.; Evans, J.L.; Groisman, P.Y.; Karl, T.R.; Kunkel, K.E.; Ambenje, P. Observed Variability and Trends in Extreme Climate Events: A Brief Review. Bull. Am. Meteorol. Soc. 2000, 81, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Tölle, M.H.; Schefczyk, L.; Gutjahr, O. Scale dependency of regional climate modeling of current and future climate extremes in Germany. Theor. Appl. Climatol. 2018, 134, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Hein, T.; Funk, A.; Pletterbauer, F.; Graf, W.; Zsuffa, I.; Haidvogl, G.; Schinegger, R.; Weigelhofer, G. Management challenges related to long-term ecological impacts, complex stressor interactions, and different assessment approaches in the Danube River Basin. River Res. Appl. 2019, 35, 500–509. [Google Scholar] [CrossRef]
- Abonyi, A.; Ács, É.; Hidas, A.; Grigorszky, I.; Várbíró, G.; Borics, G.; Kiss, K.T. Functional diversity of phytoplankton highlights long-term gradual regime shift in the middle section of the Danube River due to global warming, human impacts and oligotrophication. Freshw. Biol. 2018, 63, 456–472. [Google Scholar] [CrossRef] [Green Version]
- Soininen, J.; Jamoneau, A.; Rosebery, J.; Passy, S.I. Global patterns and drivers of species and trait composition in diatoms. Glob. Ecol. Biogeogr. 2016, 25, 940–950. [Google Scholar] [CrossRef]
- Hajósy, A. Információk a Vízlépcsőügyről és a Dunáról. [Information about the Water Dam Project and Danube]. Available online: http://www.szigetkoz.biz/ (accessed on 29 November 2019).
- Trásy, B.; Kovács, J.; Hatvani, I.G.; Havril, T.; Németh, T.; Scharek, P.; Szabó, C. Assessment of the interaction between surface- and groundwater after the diversion of the inner delta of the River Danube (Hungary) using multivariate statistics. Anthropocene 2018, 22, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Kertész, J. A vizes élőhely rehabilitáció 25 éve a Szigetközben. [25 years of rehabilitation of aquatic habitats in Szigetköz]. In Proceedings of the XXXV Országos Vándorgyűlés, Mosonmagyaróvár, Hungary, 5–7 July 2017. [Google Scholar]
- Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy. Available online: http://data.europa.eu/eli/dir/2000/60/oj/eng (accessed on 2 December 2019).
- Buczkó, K.; Ács, É. Preliminary studies on the periphytic algae in the branch-system of the Danube at Cikolasziget (Hungary). Stud. Bot. Hung. 1992, 23, 49–62. [Google Scholar]
- Ács, É.; Buczkó, K. Comparative algological studies on the periphyton in the branch-system of the River Danube at Ásványráró (Hungary). In Limnologische Berichte Donau 1994; Ambühl, H., Bloesch, J., Eds.; IAD: Dübendorf, Switzerland, 1994; pp. 413–416. [Google Scholar]
- Buczkó, K.; Rajczy, M. Changes of attached diatoms in a dead arm of the Danube between 1992–1999 at Ásványráró (Szigetköz section). Stud. Bot. Hung. 2001, 32, 39–61. [Google Scholar]
- Buczkó, K. Sixteen years of diatom monitoring of Szigetköz region of Danube—An illustrated Diatom Checklist of the Szigetköz region, Hungary. In Proceedings of the 4th Central European Diatom Meeting, Reichenau Island, Germany, 12–14 March 2010; pp. 12–14. [Google Scholar]
- CEN EN 13946. Water Quality—Guidance Standard for the Routine Sampling and Pretreatment of Benthic Diatoms from Rivers; Comité Européen de Normalisation: Geneva, Switzerland, 2003. [Google Scholar]
- Hustedt, F. Bacillariophyta. In Süßwasserflora von Mitteleuropa; Pascher, A., Ed.; G. Fischer: Jena, Germany, 1930. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 1. Teil: Naviculaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; VEB Gustav Fisher Verlag: Jena, Germany, 1986. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 2. Teil: Epithemiaceae, Bacillariaceae, Surirellaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; VEB Gustav Fisher Verlag: Jena, Germany, 1988. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 3. Teil: Eunoticeae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverzeichnis Teil 1–4. In Süßwasserflora von Mitteleuropa; Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Schmitten-Oberreifenberg, Germany, 2017. [Google Scholar]
- Coste, M. Etude des Méthodes Biologiques d’appréciation Quantitative de la Qualité des Eaux. In Rapport Division Qualité des Eaux Lyon; Agence Financière de Bassin Rhône-Méditerranée-Corse, CEMAGREF: Pierre-Bénite, France, 1982. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. “Omnidia”: Software for taxonomy, calculation of diatom indices and inventories management. Hydrobiologia 1993, 269, 509–513. [Google Scholar] [CrossRef]
- Ács, É.; Borics, G.; Kiss, K.T.; Várbíró, G. Módszertani útmutató a fitobentosz élőlénycsoport VKI szerinti gyűjtéséhez, feldolgozásához és kiértékeléséhez. In Guidance for Hungarian Phytobenthos Monitoring; 2015; Available online: http://www.kornyezetvedok.hu/vgt/vgt2/orszagos/6_1_hatteranyag_Fitobentosz_Modszertani_utmutatoVGT2.pdf?picture=pic2 (accessed on 20 November 2019).
- Rimet, F.; Bouchez, A. Life-forms, cell-sizes and ecological guilds of diatoms in European rivers. Knowl. Manag. Aquat. Ecosyst. 2012, 1283–1299. [Google Scholar] [CrossRef]
- Tapolczai, K.; Bouchez, A.; Stenger-Kovács, C.; Padisák, J.; Rimet, F. Trait-based ecological classifications for benthic algae: Review and perspectives. Hydrobiologia 2016, 776, 1–17. [Google Scholar] [CrossRef]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Netherland J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar] [CrossRef]
- Földi, A.; Ács, É.; Grigorszky, I.; Ector, L.; Wetzel, C.E.; Várbíró, G.; Kiss, K.T.; Dobosy, P.; Trábert, Z.; Borsodi, A.K.; et al. Unexpected consequences of bombing. Community level response of epiphytic diatoms to environmental stress in a saline bomb crater pond area. PLoS ONE 2018, 13, e0205343. [Google Scholar] [CrossRef]
- Marazzi, L.; Gaiser, E.E. Long-term changes in spatially structured benthic diatom assemblages in a major subtropical wetland under restoration. Inland Waters 2018, 8, 434–448. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L.F.J. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K. Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. 2016. Available online: https://CRAN.R-project.org/package=ggplot2 (accessed on 20 November 2019).
- QGIS Development Team. QGIS Geographic Information System. In Open Source Geospatial Foundation Project; Open Source Geospatial Foundation, 2019; Available online: http://qgis.osgeo.org/ (accessed on 20 November 2019).
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Poole, G.C. Fluvial landscape ecology: Addressing uniqueness within the river discontinuum. Freshw. Biol. 2002, 47, 641–660. [Google Scholar] [CrossRef] [Green Version]
- Benda, L.; Poff, N.L.; Miller, D.; Dunne, T.; Reeves, G.; Pess, G.; Pollock, M. The Network Dynamics Hypothesis: How Channel Networks Structure Riverine Habitats. BioScience 2004, 54, 413–427. [Google Scholar] [CrossRef] [Green Version]
- Tornés, E.; Cambra, J.; Gomà, J.; Leira, M.; Ortiz, R.; Sabater, S. Indicator taxa of benthic diatom communities: A case study in Mediterranean streams. Ann. Limnol. Int. J. Limnol. 2007, 43, 1–11. [Google Scholar] [CrossRef]
- Sabater, S.; Barceló, D.; De Castro-Català, N.; Ginebreda, A.; Kuzmanovic, M.; Petrovic, M.; Picó, Y.; Ponsatí, L.; Tornés, E.; Muñoz, I. Shared effects of organic microcontaminants and environmental stressors on biofilms and invertebrates in impaired rivers. Environ. Pollut. 2016, 210, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S. Mechanisms of Diatom Assembly in a Hydrologically-Managed Subtropical Wetland. Ph.D. Thesis, Florida International University, Miami, FL, USA, 2014. [Google Scholar]
- Royle, J.A.; Nichols, J.D. Estimating Abundance from Repeated Presence–Absence Data or Point Counts. Ecology 2003, 84, 777–790. [Google Scholar] [CrossRef]
- Heino, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 2013, 88, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Hanski, I. Dynamics of regional distribution: The core-satellite species hypothesis. Oikos 1982, 38, 210–221. [Google Scholar] [CrossRef]
- Keve, G. Space-Time Ice Monitoring of the Hungarian Lower-Danube. Period. Polytech. Civ. Eng. 2017, 61, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, W.; Zalewski, M. Abundance and co-occurrence patterns of core and satellite species of ground beetles on small lake islands. Oikos 2006, 114, 338–348. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Lovett-Doust, J. Core and Satellite Species in Degraded Habitats: An Analysis Using Malagasy Tree Communities. Biodivers. Conserv. 2007, 16, 2515–2529. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The Flood Pulse Concept in River-Floodplain Systems. Can. Spec. Publ. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Munteanu, N.; Maly, E.J. The effect of current on the distribution of diatoms settling on submerged glass slides. Hydrobiologia 1981, 78, 273–282. [Google Scholar] [CrossRef]
- Benenati, P.; Shannon, J.; Blinn, D. Desiccation and recolonization of phytobenthos in a regulated desert river: Colorado River at Lees Ferry, Arizona, USA. Regul. Rivers Res. Manag. 1998, 14, 519–532. [Google Scholar] [CrossRef]
- Trábert, Z.; Kiss, K.T.; Várbíró, G.; Dobosy, P.; Grigorszky, I.; Ács, É. Comparison of the utility of a frequently used diatom index (IPS) and the diatom ecological guilds in the ecological status assessment of large rivers. Fundam. Appl. Limnol. 2017, 189, 87–103. [Google Scholar] [CrossRef]
- Passy, S.I. Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquat. Bot. 2007, 86, 171–178. [Google Scholar] [CrossRef]
- Lange, K.; Liess, A.; Piggott, J.J.; Townsend, C.R.; Matthaei, C.D. Light, nutrients and grazing interact to determine stream diatom community composition and functional group structure. Freshw. Biol. 2011, 56, 264–278. [Google Scholar] [CrossRef]
- Hambrook Berkman, J.A.; Porter, S.D. An overview of algal monitoring and research in the U.S. Geological Survey’s National Water Quality Assessment (NAWQA) Program. Diatom 2004, 20, 13–22. [Google Scholar]
- Lange, K.; Townsend, C.R.; Matthaei, C.D. A trait-based framework for stream algal communities. Ecol. Evol. 2016, 6, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Ács, É.; Kiss, K.T. Effects of the water discharge on periphyton abundance and diversity in a large river (River Danube, Hungary). Hydrobiologia 1993, 249, 125–133. [Google Scholar] [CrossRef]
- Falasco, E.; Piano, E.; Bona, F. Diatom flora in Mediterranean streams: Flow intermittency threatens endangered species. Biodivers. Conserv. 2016, 25, 2965–2986. [Google Scholar] [CrossRef]
- B-Béres, V.; Tóthmérész, B.; Bácsi, I.; Borics, G.; Abonyi, A.; Tapolczai, K.; Rimet, F.; Bouchez, Á.; Várbíró, G.; Török, P. Autumn drought drives functional diversity of benthic diatom assemblages of continental intermittent streams. Adv. Water Resour. 2019, 126, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Ertl, M.; Tomajka, J. Primary production of the periphyton in the littoral of the Danube. Hydrobiologia 1973, 42, 429–444. [Google Scholar] [CrossRef]
- Rosemarin, A.S. Comparison of primary productivity (14C) per unit biomass between phytoplankton and periphyton in the Ottawa River near Ottawa, Canada. SIL Proc. 1922–2010 1975, 19, 1584–1592. [Google Scholar] [CrossRef]
- Kelly, M.G.; Cazaubon, A.; Coring, E.; Dell’Uomo, A.; Ector, L.; Goldsmith, B.; Guasch, H.; Hürlimann, J.; Jarlman, A.; Kawecka, B.; et al. Recommendations for the routine sampling of diatoms for water quality assessments in Europe. J. Appl. Phycol. 1998, 10, 215–224. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, Q.; Burford, M.A.; Sheldon, F.; Bunn, S.E. Benthic Diatom Based Indices for Water Quality Assessment in Two Subtropical Streams. Front. Microbiol. 2017, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- European Commission. CIS Guidance Document No 13 Overall Approach to the Classification of Ecological Status and Ecological Potential. In Common Implementation Strategy for the Water Framework Directive (2000/60/EC); Office for Official Publications of the European Communities: Luxemburg, 2003. [Google Scholar]
- Várbíró, G.; Borics, G.; Csányi, B.; Fehér, G.; Grigorszky, I.; Kiss, K.T.; Tóth, A.; Ács, É. Improvement of the ecological water qualification system of rivers based on the first results of the Hungarian phytobenthos surveillance monitoring. Hydrobiologia 2012, 695, 125–135. [Google Scholar] [CrossRef]
- Joint Danube Survey 3—A Comprehensive Analysis of Danube Water Quality, Final Scientific Report; Liška, I.; Wagner, F.; Sengl, M.; Deutsch, K.; Slobodník, J. (Eds.) ICPDR: Vienna, Austria, 2015. [Google Scholar]
- Pan, Y.; Stevenson, R.J.; Hill, B.H.; Kaufmann, P.R.; Herlihy, A.T. Spatial Patterns and Ecological Determinants of Benthic Algal Assemblages in Mid-Atlantic Streams, USA. J. Phycol. 1999, 35, 460–468. [Google Scholar] [CrossRef]
- Arthington, A.H.; Bunn, S.E.; Poff, N.L.; Naiman, R.J. The challenge of providing environmental flow rules to sustain river ecosystems. Ecol. Appl. Publ. Ecol. Soc. Am. 2006, 16, 1311–1318. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Palmer, M.A.; Hart, D.D.; Richter, B.D.; Arthington, A.H.; Rogers, K.H.; Meyer, J.L.; Stanford, J.A. River flows and water wars: Emerging science for environmental decision making. Front. Ecol. Environ. 2003, 1, 298–306. [Google Scholar] [CrossRef]
- ICPDR International Commission of the Danube River. Integrating and Editing New Scientific Results in Climate Change Research and the Resulting Impacts on Water Availability to Revise the Existing Adaptation Strategies in the Danube River Basin. Available online: https://www.icpdr.org/main/icpdr-climate-change-workshop-2018 (accessed on 29 November 2019).
- Nohara, D.; Kitoh, A.; Hosaka, M.; Oki, T. Impact of Climate Change on River Discharge Projected by Multimodel Ensemble. J. Hydrometeorol. 2006, 7, 1076–1089. [Google Scholar] [CrossRef] [Green Version]















© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ács, É.; Bíró, T.; Berta, C.; Duleba, M.; Földi, A.; Grigorszky, I.; Hidas, A.; Knisz, J.; Korponai, J.L.; Trábert, Z.; et al. Long-Term Changes of Species Composition and Functional Traits of Epiphytic Diatoms in the Szigetköz Region (Hungary) of the Danube River. Water 2020, 12, 776. https://doi.org/10.3390/w12030776
Ács É, Bíró T, Berta C, Duleba M, Földi A, Grigorszky I, Hidas A, Knisz J, Korponai JL, Trábert Z, et al. Long-Term Changes of Species Composition and Functional Traits of Epiphytic Diatoms in the Szigetköz Region (Hungary) of the Danube River. Water. 2020; 12(3):776. https://doi.org/10.3390/w12030776
Chicago/Turabian StyleÁcs, Éva, Tibor Bíró, Csaba Berta, Mónika Duleba, Angéla Földi, István Grigorszky, András Hidas, Judit Knisz, János L. Korponai, Zsuzsa Trábert, and et al. 2020. "Long-Term Changes of Species Composition and Functional Traits of Epiphytic Diatoms in the Szigetköz Region (Hungary) of the Danube River" Water 12, no. 3: 776. https://doi.org/10.3390/w12030776