The Relative Importance of Spatial Scale Variables for Explaning Macroinvertebrate Richness in Different Aquatic Ecological Function Regions


Abstract
:1. Introduction
2. Methods
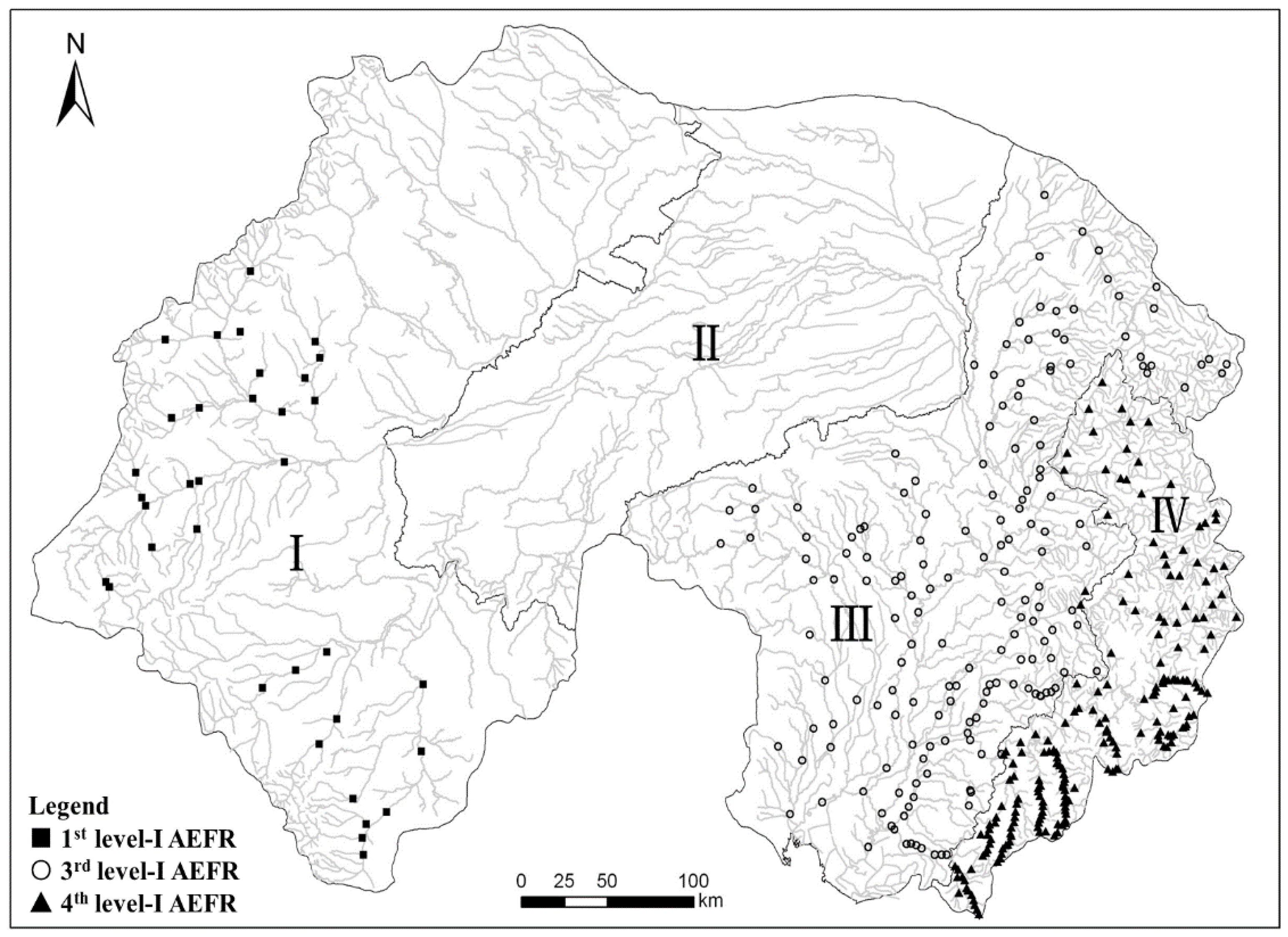
2.1. Study Area and Site Selection
2.2. Geophysical Landscape Variables
2.3. Land Use
2.4. Site Habitat Variables
2.5. Macroinvertebrate Sampling
2.6. Data Analysis
3. Results
3.1. Characteristics of Assemblage Richness and Environment Variables among the Three Level-I AEFRs
3.2. MLR Models
3.3. Relative Importance of Variables in the Combined MLR Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stiassny, M.L. An overview of freshwater biodiversity: With some lessons learned from African fishes. Fisheries 1996, 21, 7–13. [Google Scholar] [CrossRef]
- McAllister, D.E.; Hamilton, A.L.; Harvey, B. Global freshwater biodiversity: Striving for the integrity of freshwater ecosystem. Sea Wind 1997, 11, 1–140. [Google Scholar]
- Ricciardi, A.; Rasumussen, J.B. Extinction rates of North American freshwater fauna. Conserv. Biol. 1999, 13, 1220–1222. [Google Scholar] [CrossRef]
- Hughes, R.M.; Noss, R.F. Biological diversity and biological integrity: Current concerns for lakes and streams. Fisheries 1992, 17, 11–19. [Google Scholar] [CrossRef]
- Nerbonne, B.A.; Vondracek, B. Effects of local land use on physical habitat, benthic macroinvertebrate, and fish in the Whitewater River, Minnesota, USA. Environ. Manag. 2001, 28, 87–99. [Google Scholar] [CrossRef]
- Pinto, B.C.T.; Araujo, F.G.; Rodrigues, V.D.; Hughes, R.M. Local and ecoregion effects on fish assemblage structure in tributaries of the Rio Paraiba do Sul, Brazil. Freshw. Biol. 2009, 54, 2600–2615. [Google Scholar] [CrossRef]
- Macedo, D.R.; Hughes, R.M.; Ligeiro, R.; Ferreira, W.R.; Castro, M.A.; Junqueira, N.T.; Oliveira, D.R.; Firmiano, K.R.; Kaufmann, P.R.; Pompeu, P.S.; et al. The relative influence of catchment and site variables on fish and macroinvertebrate richness in cerrado biome streams. Landsc. Ecol. 2014, 29, 1001–1016. [Google Scholar] [CrossRef]
- Hilsenhoff, W.L. An improved biotic index of organic stream pollution. Great Lakes Entomol. 1987, 20, 31–39. [Google Scholar]
- Lenat, D.R. A biotic index for the southeastern United States: Derivation and list of tolerance values, with criteria for assigning water-quality rating. N. Am. Benthol. Soc. 1993, 12, 279–290. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, G.E.; Frydenborg, R.; McCarron, E.; White, J.S.; Bastian, M.L. A framework for biological criteria for Florida streams using benthic macroinvertebrates. J. N. Am. Benthol. Soc. 1996, 15, 185–211. [Google Scholar] [CrossRef]
- Dohet, A.; Ector, L.; Cauchie, H.; Hoffmann, L. Identification of benthic invertebrates and diatom indicator taxa that distinguish different stream types as well as degraded from reference conditions in Luxembourg. Anim. Biol. 2008, 58, 419–472. [Google Scholar] [CrossRef]
- USEPA. National Rivers and Stream Assessment 2008–2009: A Collaborative Survey; EPA/841/D-13/001; Office of Wetlands, Oceans and Watersheds and Office of Research and Development: Washington, DC, USA, 2013.
- Chen, K.; Hughes, R.M.; Xu, S.; Zhang, J.; Cai, D.; Wang, B. Evaluating performance of macroinvertebrate-based predictive and null modeled multimetric indices (MMI) using multi-season and multi-year samples. Ecol. Indic. 2014, 36, 142–151. [Google Scholar] [CrossRef]
- Brown, L.R.; Cuffney, T.F.; Coles, J.F.; Fitzpatrick, F.; McMahon, G.; Steuer, J.; Bell, A.H.; May, J.T. Urban streams across the USA: Lessons learned from studies in 9 metropolitan areas. J. N. Am. Benthol. Soc. 2009, 28, 1051–1069. [Google Scholar] [CrossRef]
- Frissell, C.A.; Liss, W.J.; Warren, C.E.; Hurley, M.D. A hierarchical framework for stream habitat classification: Viewing streams in a watershed context. Environ. Manag. 1986, 10, 199–214. [Google Scholar] [CrossRef]
- Strayer, D.L.; Beighley, R.E.; Thompson, L.C.; Brooks, S.; Nilsson, C.; Pinay, G.; Naiman, R.J. Effects of land cover on stream ecosystems: Roles of empirical models and scaling issues. Ecosystems 2003, 6, 407–423. [Google Scholar] [CrossRef]
- Townsend, C.R.; Doledec, S.; Norris, R.; Peacock, K.; Arbuckle, C. The influence of scale and geography on relationships between stream community composition and landscape variables: Description and prediction. Freshw. Biol. 2003, 48, 768–785. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Ann. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef]
- Allan, J.D.; Erickson, D.L.; Fay, J. The influence of catchment land use on stream integrity across multiple spatial scales. Freshw. Biol. 1997, 37, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Bledsoe, B.P.; Watson, C.C. Effects of urbanization on channel instability. J. Am. Water Resour. Assoc. 2001, 37, 255–270. [Google Scholar] [CrossRef]
- Bojsen, B.H.; Barriga, R. Effects of deforestation on fish community structure in Ecuadorian Amazon streams. Freshw. Biol. 2002, 47, 2246–2260. [Google Scholar] [CrossRef]
- Weijters, M.J.; Janse, J.H.; Alkemade, R.; Verhoeven, J.T.A. Quantifying the effect of catchment land use and water nutrient concentrations on freshwater river and stream biodiversity. Aquat. Conserv. Mar. Freshw. Ecosyst. 2009, 19, 104–112. [Google Scholar] [CrossRef]
- Nilsson, M.; Persson, A. Can Earth system interactions be governed? Governance functions for linking climate change mitigation with land use, freshwater and biodiversity protection. Ecol. Econ. 2012, 75, 61–71. [Google Scholar] [CrossRef]
- Ferreira, W.R.; Ligeiro, R.; Macedo, D.R.; Hughes, R.M.; Kaufmann, P.R.; Oliveira, L.G.; Callisto, M. Importance of environmental factors for the richness and distribution of benthic macroinvertebrates in tropical headwater streams. Freshw. Sci. 2014, 33, 860–871. [Google Scholar] [CrossRef]
- Wang, H.; Qin, D.Y.; Wang, J.H. The study on multi-scale regional water cycle simulation and two-terms water cycle mode. In Evolution Law and Maintaining Mechanism of Renewable Capability of Water Resources in the Yellow River Basin; Liu, C.M., Chen, X.G., Eds.; Publishing House for Yellow Water Conservancy: Zhengzhou, China, 2001; pp. 34–42. (In Chinese) [Google Scholar]
- Stauffer, J.C.; Goldstein, R.M.; Newman, R.M. Relationship of wooded riparian zones and runoff potential to fish community composition in agricultural streams. Can. J. Fish. Aquat. Sci. 2000, 57, 307–316. [Google Scholar] [CrossRef]
- Johnson, L.B.; Breneman, D.H.; Richards, C. Macroinvertebrate community structure and function associated with large wood in low gradient streams. River Res. Appl. 2003, 19, 199–218. [Google Scholar] [CrossRef]
- Kolpin, D.W.; Furlong, E.T.; Meyer, M.T.; Thurman, E.M.; Zaugg, S.D.; Barber, L.B.; Buxton, H.T. Pharmaceuticals, hormones, and other organic wastewater contaminants in US streams, 1999–2000: A national reconnaissance. Environ. Sci. Technol. 2002, 36, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.M.; Stroud, M.J. Water quality and sediment and nutrient export from New Zealand hill-land catchments of contrasting land use. N. Z. J. Mar. Freshw. Res. 2002, 36, 409–429. [Google Scholar] [CrossRef]
- Niyogi, D.K.; Simon, K.S.; Townsend, C.R. Land use and stream ecosystem functioning: Nutrient uptake in streams that contrast in agricultural development. Arch. Hydrobiol. 2004, 160, 471–486. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Stanley, E.H.; Vander Zanden, M.J. State of the World’s Freshwater Ecosystems: Physical, Chemical, and Biological Changes. Ann. Rev. Environ. Resour. 2011, 36, 75–99. [Google Scholar] [CrossRef]
- Saly, P.; Takacs, P.; Kiss, L.; Bíró, P.; Eros, T. The relative influence of spatial context and catchment- and site-scale environmental factors on stream fish assemblages in a human-modified landscape. Ecol. Freshw. Fish 2011, 20, 251–262. [Google Scholar] [CrossRef]
- Marzin, A.; Verdonschot, P.F.M.; Pont, D. The relative influence of catchment, riparian corridor, and reach-scale anthropogenic pressures on fish and macroinvertebrate assemblages in French rivers. Hydrobiologia 2012, 704, 375–388. [Google Scholar] [CrossRef]
- Omernik, J. Map supplement: Ecoregions of the conterminous United States. Ann. Assoc. Am. Geogr. 1987, 77, 118–125. [Google Scholar] [CrossRef]
- Biggs, B.; Duncan, M.; Jowett, I.; Quinn, J.; Hickey, C.; Davies-Colley, R.; Close, M. Ecological characterization, classification, and modelling of New Zealand rivers: An introduction and synthesis. N. Z. J. Mar. Freshw. Res. 1990, 24, 277–304. [Google Scholar] [CrossRef]
- Harding, J.S.; Winterbourn, M.J. New Zealand Ecoregions: A Classification for Use in Stream Conservation and Management; Department of Conservation: Wellington, New Zealand, 1997.
- Wells, F.; Newall, P. An Examination of an Aquatic Ecoregion Protocol for Australia; Australian and New Zealand Environment and Conservation Council: Canberra, Australia, 1997.
- Kleynhans, C.; Silberbauer, M.; Kemper, N. Preliminary Ecoregion Level 1 Classification for South Africa; Institute for Water Quality Studies; Department of Water Affairs and Forestry and Institute for Water Research: Pretoria, South Africa, 1998.
- EU Water Framework Directive. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for Community action in the field of water policy. Off. J. Eur. Communities 2000, L327, 1–72. [Google Scholar]
- Ding, S.; Zhang, Y.; Liu, B.; Kong, W.J.; Meng, W. Effects of riparian land use on water quality and fish communities in the headwater stream of the Taizi River in China. Front. Environ. Sci. Eng. 2013, 7, 699–708. [Google Scholar] [CrossRef]
- Gao, X.; Niu, C.J.; Chen, Y.S.; Yin, X.W. Spatial heterogeneity of stream environmental conditions and macroinvertebrates community in an agriculture dominated watershed and management implications for a large river (the Liao River, China) basin. Environ. Monit. Assess. 2014, 186, 2375–2391. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, Y.; Ding, S.; Zhao, R.; Meng, W. Response of fish communities to environmental changes in an agriculturally dominated watershed (Liao River Basin) in northeastern China. Ecol. Eng. 2015, 75, 130–141. [Google Scholar] [CrossRef]
- Li, Y.L.; Liu, K.; Li, L.; Xu, Z.X. Relationship of land use/cover on water quality in the Liao River basin, China. Procedia Environ. Sci. 2012, 13, 1484–1493. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.M.; Xiong, J.; Qiu, J.W.; Wu, J.M.; Wang, J.W.; Xie, Z.C. Structure of Macroinvertebrate Community in Relation to environmental parameters in a Subtropical Asian River System. Int. Rev. Hydrobiol. 2010, 95, 42–57. [Google Scholar] [CrossRef]
- Astorga, A.; Heino, J.; Luoto, M.; Muotka, T. Freshwater biodiversity at regional extent: Determinants of macroinvertebrate taxonomic richness in headwater streams. Ecogrphy 2011, 34, 705–713. [Google Scholar] [CrossRef]
- Hermoso, V.; Linke, S.; Prenda, J. Identifying priority sites for the conservation of freshwater fish biodiversity in a Mediterranean basin with a high degree of threatened endemics. Hydrobiologia 2009, 623, 127–140. [Google Scholar] [CrossRef]
- Marchant, R.; Hirst, A.; Norris, R.; Metzeling, L. Classification of macroinvertebrate communities across drainage basins in Victoria, Australia: Consequences of sampling on a broad spatial scale for predictive modeling. Freshw. Biol. 1999, 41, 253–268. [Google Scholar] [CrossRef]
- Marchant, R. Do rare species have any place in multivariate analysis for bioassessment? J. N. Am. Benthol. Soc. 2002, 21, 311–313. [Google Scholar] [CrossRef]
- Goldstein, R.M.; Carlisle, D.M.; Meador, M.R.; Short, T.M. Can basin land use effects on physical characteristics of streams be determined at broad geographic scales? Environ. Monit. Assess. 2007, 130, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.; Moss, D.; Armitage, P.; Furse, M. A preliminary classification of running-water sites in Great Britain based on macro-invertebrate species and the prediction of community type using environmental data. Freshw. Biol. 1984, 14, 221–256. [Google Scholar] [CrossRef]
- Holmes, N.; Boon, P.; Rowell, T. A revised classification system for British rivers based on their aquatic plant communities. Aquat. Conserv. Mar. Freshw. Ecosyst. 1998, 8, 555–578. [Google Scholar] [CrossRef]
- Foerster, J.; Gutowski, A.; Schaumburg, J. Defining types of running waters in Germany using benthic algae: A prerequisite for monitoring according to the Water Framework Directive. J. Appl. Phycol. 2004, 16, 407–418. [Google Scholar] [CrossRef]
- Lorenz, A.; Feld, C.; Hering, D. Typology of streams in Germany based on benthic invertebrates: Ecoregions, zonation, geology and substrate. Limnologica 2004, 34, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Tison, J.; Park, Y.S.; Coste, M.; Wasson, J.G.; Ector, L.; Rimet, F.; Delmas, F. Typology of diatom communities and the influence of hydro-ecoregions: A study on the French hydrosystem scale. Water Res. 2005, 39, 3177–3188. [Google Scholar] [CrossRef]
- Di Lascio, A.; Rossi, L.; Carlino, P.; Calizza, E.; Rossi, D.; Costantini, M.L. Stable isotope variation in macroinvertebrates indicates anthropogenic disturbance along an urban stretch of the river Tiber (Rome, Itlay). Ecol. Indic. 2013, 28, 107–114. [Google Scholar] [CrossRef]
- Wang, R.Z.; Xu, T.L.; Yu, L.Z.; Zhu, J.J.; Li, X.Y. Effects of land use types on surface water quality across an anthropogenic disturbance gradient in the upper reach of the Hun River, Northeast China. Environ. Monit. Assess. 2013, 185, 4141–4151. [Google Scholar] [CrossRef] [PubMed]
- Kay, W.R.; Halse, S.A.; Scanlon, M.D.; Smith, S.J. Distribution and environmental tolerances of aquatic macroinvertebrate families in the agricultural zone of southwestern Australia. J. N. Am. Benthol. Soc. 2001, 20, 182–199. [Google Scholar] [CrossRef]
- De Deckker, P. Australian salt lakes: Their history, chemistry, and biota—A review. Hydrobiologia 1983, 105, 231–244. [Google Scholar] [CrossRef]
- Graf, W.L. Dam nation: Geographic census of American dams and their larger scale hydrological impacts. Water Resour. Res. 1999, 35, l305–l310. [Google Scholar] [CrossRef]
- Bunn, S.E.; Arthington, A.H. Basic Principles and Ecological Consequences of Altered Flow Regimes for Aquatic Biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef] [Green Version]
- Kurunc, A.; Yurekli, K.; Okman, C. Effects of Kilickaya Dam on concentration and load values of water quality constituents in Kelkit Stream in Turkey. J. Hydrol. 2006, 317, 17–30. [Google Scholar] [CrossRef]
- Li, R.N.; Chen, Q.; Tonina, D.; Cai, D. Effects of upstream reservoir regulation on the hydrological regime and fish habitats of the Lijiang River, China. Ecol. Eng. 2015, 76, 75–83. [Google Scholar] [CrossRef]
- Beisel, J.N.; Usseglio-Polatera, P.; Thomas, S.; Moreteau, J. Stream community structure in relation to spatial variation: The influence of mesohabitat characteristics. Hydrobiologia 1998, 389, 73–88. [Google Scholar] [CrossRef]
- Jones, J.I.; Murphy, J.F.; Collins, A.L.; Sear, D.A.; Naden, P.S.; Armitage, P.D. The impact of fine sediment on macro-invertebrates. River Res. Appl. 2012, 28, 1055–1071. [Google Scholar] [CrossRef]
- Kaufmann, P.R.; Larsen, D.P.; Faustini, J.M. Bed stability and sedimentation associated with human disturbances in Pacific Northwest streams. J. Am. Water Resour. Assoc. 2009, 45, 434–459. [Google Scholar] [CrossRef]
- Culp, J.M.; Walde, S.J.; Davies, R.W. Relative importance of substrate particle size and detritus to stream benthic macroinvertebrate microdistribution. Can. J. Fish. Aquat. Sci. 1983, 40, 1568–1574. [Google Scholar] [CrossRef]
- Peckarsky, B.L. Habitat selection by stream-dwelling predatory stoneflies. Can. J. Fish. Aquat. Sci. 1991, 48, 1069–1076. [Google Scholar] [CrossRef]
- Sarriquet, P.E.; Bordenave, P.; Marmonier, P. Effects of bottom sediment restoration on interstitial habitat characteristics and benthic macroinvertebrate assemblages in a headwater stream. River Res. Appl. 2007, 23, 815–828. [Google Scholar] [CrossRef]
- Culp, J.M.; Wrona, F.J.; Davies, R.W. Response of stream benthos and drift to fine sediment deposition versus transport. Can. J. Zool. 1986, 64, 1345–1351. [Google Scholar] [CrossRef]
- Armitage, P.D.; Cannan, C.E. Annual changes in summer patterns of mesohabitat distribution and associated macroinvertebrate assemblages. Hydrol. Process. 2000, 14, 3161–3179. [Google Scholar] [CrossRef]
- Couceiro, S.R.M.; Hamada, N.; Luz, S.L.B.; Forsberg, B.R.; Pimentel, T.P. Deforestation and sewage effects on aquatic macroinvertebrates in urban streams in Manaus, Amazonas, Brazil. Hydrobiologia 2007, 575, 271–284. [Google Scholar] [CrossRef]
- Deegan, L.A.; Peterson, B.J.; Golden, H.; McIvor, C.C.; Miller, M.C. Effects of fish density and river fertilization on algal standing stocks, invertebrate communities, and fish production in an arctic river. Can. J. Fish. Aquat. Sci. 1997, 54, 269–283. [Google Scholar] [CrossRef]
- Perrin, C.J.; Richardson, J.S. N and P limitation of benthos abundance in the Nechako River, British Columbia. Can. J. Fish. Aquat. Sci. 1997, 54, 2574–2583. [Google Scholar] [CrossRef]
- Miltner, R.J.; Rankin, E.T. Primary nutrients and the biotic integrity of rivers and streams. Freshw. Biol. 1998, 40, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Hart, D.D.; Robinson, C.T. Resource limitation in a stream community: Phosphorus enrichment effects on periphyton and grazers. Ecology 1990, 71, 1494–1502. [Google Scholar] [CrossRef]
- Lenat, D.R.; Crawford, J.K. Effects of land use on water quality and aquatic biota of three North Carolina Piedmont streams. Hydrobiologia 1994, 294, 185–199. [Google Scholar] [CrossRef]
- Wang, L.; Lyons, J.; Kanehl, P.; Gatti, R. Influences of watershed land use on habitat quality and biotic integrity in Wisconsin streams. Fisheries 1997, 22, 6–12. [Google Scholar] [CrossRef]
- Genito, D.; Gburek, W.J.; Sharpley, A.N. Response of stream macroinvertebrates to agricultural land cover in a small watershed. J. Freshw. Ecol. 2002, 17, 109–119. [Google Scholar] [CrossRef]
- Cooper, C.M. Biological effects of agriculturally derived surface water pollutants on aquatic systems—A review. J. Environ. Qual. 1993, 22, 402–408. [Google Scholar] [CrossRef]
- Johnson, L.B.; Richards, C.; Host, G.E.; Arthur, J.W. Landscape influences on water chemistry in Midwestern stream ecosystems. Freshw. Biol. 1997, 37, 193–208. [Google Scholar] [CrossRef]
- Delong, M.D.; Brusven, M.A. Macroinvertebrate communitys structure along the longitudinal gradient of an agriculturally impacted stream. Environ. Manag. 1998, 22, 445–457. [Google Scholar] [CrossRef]
- Sponseller, R.A.; Benfield, E.F.; Valett, H.M. Relationships between land use, spatial scale and stream macroinvertebrate communities. Freshw. Biol. 2001, 46, 1409–1424. [Google Scholar] [CrossRef]


{kind=link}
| Scale | Level | Variable | Method | Code |
|---|---|---|---|---|
| Catchment | Geophysical landscape | Longitude | in situ | Longitude |
| Latitude | in situ | Latitude | ||
| Altitude | in situ | Altitude | ||
| Slope | GIS | Slope | ||
| Sinuosity | GIS | Sinuosity | ||
| Stream Order | GIS | Streamorder | ||
| Reach Length | GIS | Length | ||
| Distance from Mouth | GIS | DisfrMouth | ||
| Average Temperature | GIS | AverTem | ||
| Average Rainfall | GIS | AverRain | ||
| Up-catchment Area | GIS | UpcatchArea | ||
| Land use | Un-used Area | GIS | UnusedArea | |
| Water Area | GIS | WaterArea | ||
| Forest Area | GIS | ForestArea | ||
| Constructed Area | GIS | ConstrArea | ||
| Crop Area | GIS | CropArea | ||
| Grass Area | GIS | GrassArea | ||
| Site | Site habitat | Depth | in situ | Depth |
| Velocity | in situ | Velocity | ||
| Boulder | in situ | Boulder | ||
| Cobble | in situ | Cobble | ||
| Pebble | in situ | Pebble | ||
| Gravel | in situ | Gravel | ||
| Sand/Silt | in situ | Sand/Silt | ||
| Water Temperature | in situ | WaterTem | ||
| Electric conductivity | in situ | EC | ||
| Total Dissolved Solids | in situ | TDS | ||
| pH | in situ | pH | ||
| Dissolved Oxygen | in situ | DO | ||
| Total Nitrogen | in situ & lab | TN | ||
| Total Phosphorus | in situ & lab | TP | ||
| Ammonia Nitrogen | in situ & lab | NH3-N |
| Variables | AEFRs | p-Value | ||
|---|---|---|---|---|
| 1st | 3rd | 4th | ||
| Assemblage Richness | 10.90 ± 4.36 | 10.68 ± 5.77 | 17.32 ± 11.95 | <0.01 a,c |
| Altitude | 816.50 ± 228.58 | 91.84 ± 75.18 | 265.84 ± 125.60 | <0.01 a,c |
| Slope | 5.78 ± 4.24 | 2.19 ± 3.56 | 8.22 ± 7.85 | <0.01 a,c |
| Sinuosity | 1.16 ± 0.11 | 1.30 ± 0.22 | 1.32 ± 0.32 | <0.01 a,c |
| Streamorder | 3.00 ± 1.11 | 3.07 ± 1.48 | 2.35 ± 1.15 | <0.01 a,b |
| Length | 11.38 ± 10.00 | 12.84 ± 13.00 | 12.81 ± 11.26 | 0.531 b |
| DisfrMouth | 119.32 × 104 ± 4.91 × 104 | 33.70 × 104 ± 19.94 × 104 | 37.00 × 104 ± 9.79 × 104 | <0.01 a,c |
| AverTem | 4.10 ± 2.08 | 7.64 ± 1.11 | 5.78 ± 1.52 | <0.01 a,c |
| AverRain | 443.09 ± 71.95 | 645.63 ± 72.96 | 867.74 ± 73.85 | <0.01 b |
| UpcatchArea | 20.24 × 108 ± 25.26 × 108 | 147.03 × 108 ± 450.11 × 108 | 5.92 × 108 ± 10.88 × 108 | <0.01 a,c |
| UnusedArea | 8.99 ± 11.28 | 1.11 ± 6.02 | 0.16 ± 0.87 | <0.01 a,c |
| WaterArea | 4.04 ± 4.26 | 8.34 ± 12.64 | 2.05 ± 4.00 | <0.01 a,c |
| ForestArea | 7.78 ± 10.68 | 11.22 ± 16.96 | 66.98 ± 16.99 | <0.01 a,c |
| ConstrArea | 4.47 ± 5.02 | 10.33 ± 8.50 | 2.67 ± 3.58 | <0.01 a,c |
| CropArea | 31.16 ± 25.83 | 67.22 ± 19.42 | 26.93 ± 13.75 | <0.01 a,c |
| GrassArea | 43.56 ± 25.59 | 1.78 ± 4.60 | 1.21 ± 2.17 | <0.01 a,c |
| Depth | 19.69 ± 13.74 | 30.78 ± 17.14 | 22.56 ± 10.15 | <0.01 a,c |
| Velocity | 1.51 ± 0.96 | 0.54 ± 0.62 | 0.51 ± 0.39 | <0.01 a,c |
| Boulder | 20.26 ± 24.71 | 9.99 ± 19.81 | 39.44 ± 23.41 | <0.01 a,c |
| Cobble | 12.61 ± 15.83 | 7.14 ± 12.10 | 19.32 ± 10.86 | <0.01 a,c |
| Pebble | 15.12 ± 17.21 | 10.52 ± 15.64 | 23.26 ± 13.31 | <0.01 a,c |
| Gravel | 6.52 ± 10.93 | 5.74 ± 9.43 | 9.66 ± 8.25 | <0.01 b |
| Sand/Silt | 45.49 ± 47.27 | 62.46 ± 42.79 | 8.33 ± 14.77 | <0.01 a,c |
| WaterTem | 20.58 ± 4.56 | 20.44 ± 3.42 | 17.11 ± 4.73 | <0.01 a,c |
| EC | 373.54 ± 109.02 | 494.61 ± 181.18 | 233.25 ± 154.66 | <0.01 b |
| TDS | 180.14 ± 53.23 | 309.62 ± 129.27 | 161.17 ± 106.19 | <0.01 a,c |
| Ph | 8.35 ± 0.37 | 8.13 ± 0.52 | 8.46 ± 0.59 | <0.01 b |
| DO | 8.02 ± 1.56 | 8.53 ± 2.42 | 11.19 ± 3.85 | <0.01 a,b |
| TN | 2.62 ± 1.83 | 3.96 ± 5.18 | 3.45 ± 2.91 | <0.05 a,c |
| TP | 0.11 ± 0.14 | 0.24 ± 0.38 | 0.11 ± 0.18 | <0.01 a,c |
| NH3-N | 0.67 ± 0.95 | 1.10 ± 2.56 | 0.46 ± 0.69 | 0.065 a,c |
| AEFR | Variable | β | Std-Error | R2 | D-W Value |
|---|---|---|---|---|---|
| 1st | Geophysical landscape | 0.602 ** | 2.339 | ||
| DisfrMouth | 5.220 | 1.204 | |||
| UpcatchArea | −0.117 | 0.041 | |||
| Land use | 0.503 ** | 1.971 | |||
| UnusedArea | −0.412 | 0.094 | |||
| WaterArea | −0.573 | 0.158 | |||
| Site habitat | 0.505 ** | 1.876 | |||
| TP | −1.154 | 0.525 | |||
| TN | 0.211 | 0.112 | |||
| Depth | −0.153 | 0.078 | |||
| Sand/Silt | −0.056 | 0.038 | |||
| 3rd | Geophysical landscape | 0.112 ** | 1.588 | ||
| AverRain | −1.502 | 0.431 | |||
| UpcatchArea | −0.062 | 0.022 | |||
| Length | −0.088 | 0.058 | |||
| Land use | 0.141 ** | 1.472 | |||
| WaterArea | −0.315 | 0.089 | |||
| ConstrArea | −0.459 | 0.160 | |||
| UnusedArea | −0.330 | 0.185 | |||
| Site habitat | 0.246 ** | 1.843 | |||
| NH3–N | −0.173 | 0.122 | |||
| Boulder | 0.115 | 0.078 | |||
| TN | −0.174 | 0.073 | |||
| Sand/Silt | −0.088 | 0.041 | |||
| 4th | Geophysical landscape | 0.302 ** | 1.206 | ||
| AverRain | 1.108 | 0.528 | |||
| AverTem | −0.882 | 0.166 | |||
| Latitude | −16.851 | 3.918 | |||
| Length | −0.083 | 0.046 | |||
| Land use | 0.065 ** | 0.847 | |||
| ForestArea | 0.355 | 0.094 | |||
| Site habitat | 0.395 ** | 1.517 | |||
| Sand/Silt | −0.390 | 0.079 | |||
| Velocity | −0.906 | 0.164 | |||
| TN | −0.256 | 0.055 | |||
| WaterTem | 0.417 | 0.115 | |||
| TP | −0.527 | 0.266 | |||
| Boulder | 0.111 | 0.061 |
| AEFR | Geophysical Landscape (GL) | Land Use (LU) | Site Habitat (SH) | Total R2 | Shared Explained Variance | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | Std-Error | R2 | β | Std-Error | R2 | β | Std-Error | R2 | GL + LU R2 | GL + SH R2 | LU + SH R2 | Combined R2 | |||||
| 1st | 0.123 | 0.046 | 0.011 | 0.745 | 0.371 | 0.408 | 0.386 | 0.300 | |||||||||
| DisfrMouth | 4.461 | 1.151 | CropArea | 0.170 | 0.078 | TP | −0.925 | 0.419 | |||||||||
| AverTem | −0.174 | 0.105 | Depth | −0.171 | 0.065 | ||||||||||||
| 3rd | 0.054 | 0.111 | 0.121 | 0.378 | −0.004 | 0.091 | 0.063 | 0.029 | |||||||||
| AverRain | −1.088 | 0.499 | WaterArea | 0.009 | 0.116 | NH3-N | 0.056 | 0.138 | |||||||||
| Sinuosity | −0.871 | 0.510 | ForestArea | 0.541 | 0.123 | Boulder | 0.111 | 0.065 | |||||||||
| DisfrMouth | −0.119 | 0.080 | CropArea | 0.431 | 0.149 | TN | −0.213 | 0.089 | |||||||||
| pH | −2.236 | 0.912 | |||||||||||||||
| DO | 0.242 | 0.147 | |||||||||||||||
| 4th | 0.104 | 0.245 | 0.547 | 0.065 | 0.150 | 0.017 | 0.017 | ||||||||||
| AverRain | 0.584 | 0.463 | Sand/Silt | −0.258 | 0.071 | ||||||||||||
| Latitude | −17.835 | 4.598 | TN | −0.277 | 0.061 | ||||||||||||
| AverTem | −0.830 | 0.150 | WaterTem | 0.527 | 0.113 | ||||||||||||
| Slop | 0.059 | 0.041 | Velocity | −0.442 | 0.180 | ||||||||||||
| Boulder | 0.095 | 0.055 | |||||||||||||||
| Depth | 0.169 | 0.078 | |||||||||||||||
| NH3-N | −0.207 | 0.111 | |||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Jia, X.; Lin, J.; Qian, C.; Gao, X.; Ding, S. The Relative Importance of Spatial Scale Variables for Explaning Macroinvertebrate Richness in Different Aquatic Ecological Function Regions. Water 2019, 11, 1550. https://doi.org/10.3390/w11081550
Zhang Y, Jia X, Lin J, Qian C, Gao X, Ding S. The Relative Importance of Spatial Scale Variables for Explaning Macroinvertebrate Richness in Different Aquatic Ecological Function Regions. Water. 2019; 11(8):1550. https://doi.org/10.3390/w11081550
Chicago/Turabian StyleZhang, Yuan, Xiaobo Jia, Jianing Lin, Chang Qian, Xin Gao, and Sen Ding. 2019. "The Relative Importance of Spatial Scale Variables for Explaning Macroinvertebrate Richness in Different Aquatic Ecological Function Regions" Water 11, no. 8: 1550. https://doi.org/10.3390/w11081550