Carbon Transfer from Cyanobacteria to Pelagic and Benthic Consumers in a Subtropical Lake: Evidence from a 13C Labelling Experiment

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Detritus Preparation and Labelling
2.3. Labelling Experiment
2.4. Sample Collection
2.5. Stable Isotope Analysis
2.6. Data and Statistical Analyses
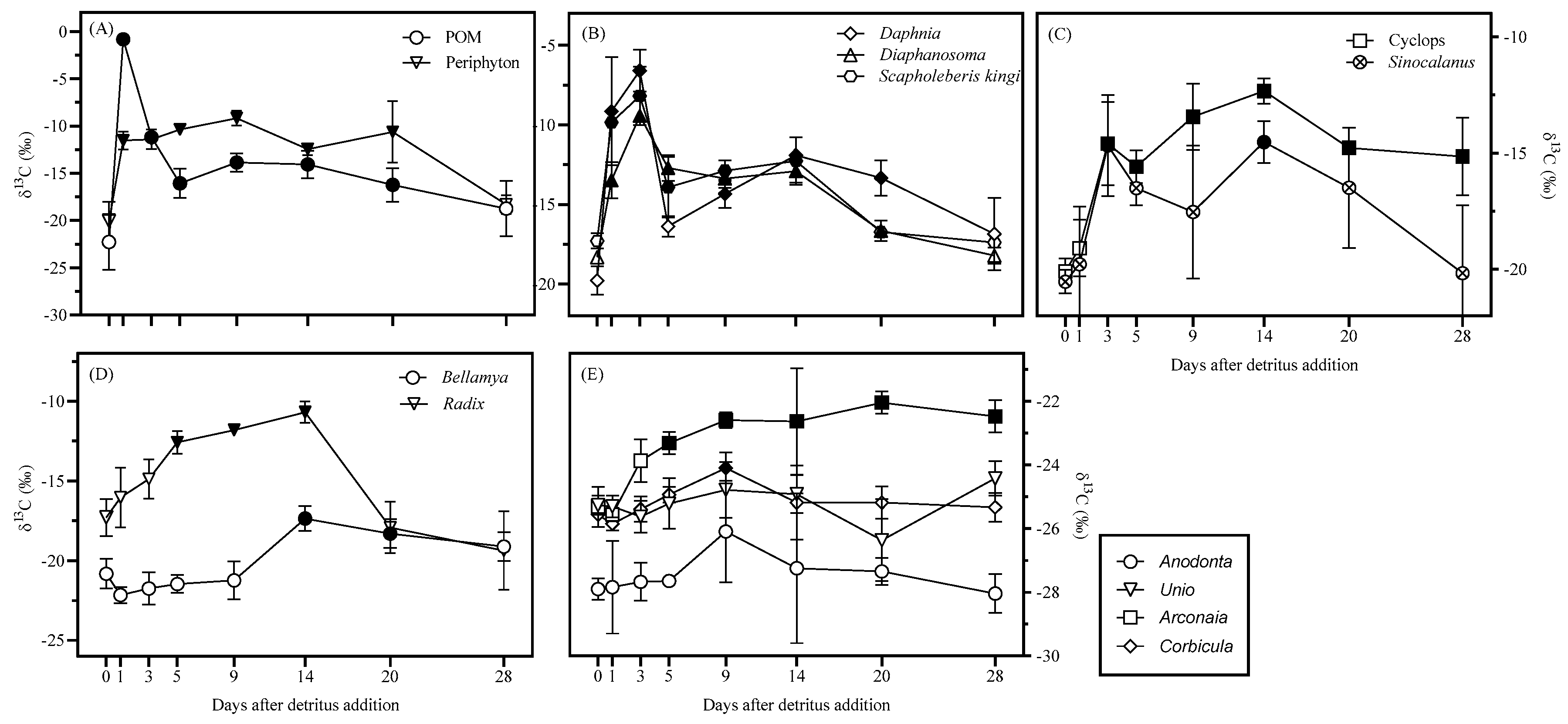
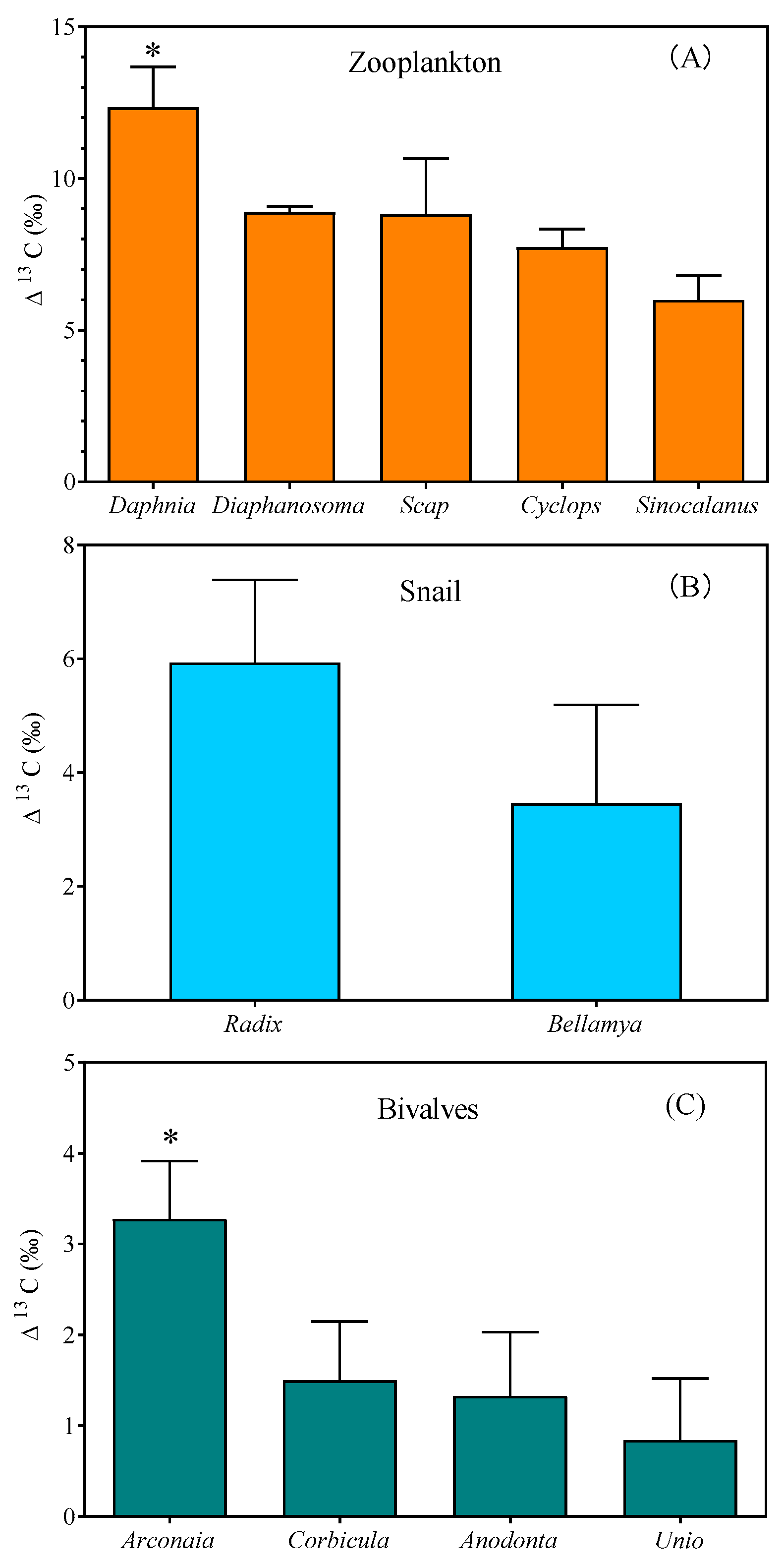
3. Results
4. Discussions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moss, B. Engineering and biological approaches to the restoration from eutrophication of shallow lakes in which aquatic plant communities are important components. Hydrobiologia 1990, 200, 367–378. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.-L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- McQueen, D.J.; Johannes, M.R.S.; Post, J.R.; Stewart, T.J.; Lean, D.R.S. Bottom-up and top-down impacts on freshwater pelagic community structure. Ecol. Monogr. 1989, 59, 289–309. [Google Scholar] [CrossRef]
- Hairston, N.G., Jr.; Hairston, N.G., Sr. Cause-effect relationships in energy flow, trophic structure, and interspecific interactions. Am. Nat. 1993, 142, 379–411. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Turner, A.M.; Hall, D.J.; Rettig, J.E.; Osenberg, C.W. Perturbation and resilience: A long-term, whole-lake study of predator extinction and reintroduction. Ecology 1995, 76, 2347–2360. [Google Scholar] [CrossRef]
- Lindeman, R.L. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–418. [Google Scholar] [CrossRef]
- Hecky, R.E.; Hesslein, R.H. Contributions of benthic algae to lake food webs as revealed by stable isotope analysis. J. N. Am. Benthol. Soc. 1995, 14, 631–653. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Vadeboncoeur, Y. Fishes as integrators of benthic and pelagic food webs in lakes. Ecology 2002, 83, 2152–2161. [Google Scholar] [CrossRef]
- Genkai-Kato, M.; Vadeboncoeur, Y.; Liboriussen, L.; Jeppesen, E. Benthic-planktonic coupling, regime shifts, and whole-lake primary production in shallow lakes. Ecology 2012, 93, 619–631. [Google Scholar] [CrossRef]
- Zhang, P.; Tang, H.; Gong, Z.; Xie, P.; Xu, J. Phytoplankton abundance constrains planktonic energy subsidy to benthic food web. J. Ecosyst. Ecogr. 2013, 4, 139. [Google Scholar]
- Zhang, X.; Liu, Z.; Jeppesen, E.; Taylor, W.D. Effects of deposit-feeding tubificid worms and filter-feeding bivalves on benthic-pelagic coupling: Implications for the restoration of eutrophic shallow lakes. Water Res. 2014, 50, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Wylie, J.L.; Currie, D.J. The relative importance of bacteria and algae as food sources for crustacean zooplankton. Limnol. Oceanogr. 1991, 36, 708–728. [Google Scholar] [CrossRef]
- Brett, M.T.; Kainz, M.J.; Taipale, S.J.; Seshan, H. Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production. Proc. Natl. Acad. Sci. USA 2009, 106, 21197–21201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautio, M.; Mariash, H.; Forsström, L. Seasonal shifts between autochthonous and allochthonous carbon contributions to zooplankton diets in a subarctic lake. Limnol. Oceanogr. 2011, 56, 1513–1524. [Google Scholar] [CrossRef] [Green Version]
- Dionisio Pires, L.M.; Bontes, B.M.; van Donk, E.; Ibelings, B.W. Grazing on colonial and filamentous toxic and non-toxic cyanobacteria by the zebra mussel Dreissena polymorpha. J. Plankton Res. 2005, 27, 331–339. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, P.; Wu, X.-P. Grazing on toxic and non-toxic Microcystis aeruginosa PCC7820 by Unio douglasiae and Corbicula fluminea. Limnology 2009, 10, 1–5. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Johengen, T.H.; Liebig, A.R. Feedback between zebra mussel selective feeding and algal composition affects mussel condition: Did the regime changer pay a price for its success? Freshw. Biol. 2009, 54, 47–63. [Google Scholar] [CrossRef]
- Ke, Z.; Xie, P.; Guo, L.; Liu, Y.; Yang, H. In situ study on the control of toxic Microcystis blooms using phytoplanktivorous fish in the subtropical Lake Taihu of China: A large fish pen experiment. Aquaculture 2007, 265, 127–138. [Google Scholar] [CrossRef]
- Zhou, Q.; Xie, P.; Xu, J.; Ke, Z.; Guo, L. Growth and food availability of silver and bighead carps: Evidence from stable isotope and gut content analysis. Aquacult. Res. 2009, 40, 1616–1635. [Google Scholar] [CrossRef]
- Guo, L.; Wang, Q.; Xie, P.; Tao, M.; Zhang, J.; Niu, Y.; Ma, Z. A non-classical biomanipulation experiment in Gonghu Bay of Lake Taihu: Control of Microcystis blooms using silver and bighead carp. Aquac. Res. 2015, 46, 2211–2224. [Google Scholar] [CrossRef]
- Watson, S.B.; McCauley, E.; Downing, J.A. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol. Oceanogr. 1997, 42, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S. What factors influence the species composition of phytoplankton in lakes of different trophic status? Hydrobiologia 1998, 369, 11–26. [Google Scholar] [CrossRef]
- Chen, Y.; Qin, B.; Teubner, K.; Dokulil, M.T. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J. Plankton Res. 2003, 25, 445–453. [Google Scholar] [CrossRef]
- Song, X.; Liu, Z.; Yang, G.; Chen, Y. Effects of resuspension and eutrophication level on summer phytoplankton dynamics in two hypertrophic areas of Lake Taihu, China. Aquat. Ecol. 2010, 44, 41–54. [Google Scholar] [CrossRef]
- Lindholm, T.; Eriksson, J.E.; Meriluoto, J.A.O. Toxic cyanobacteria and water quality problems—Examples from a eutrophic lake on Åland, South West Finland. Water Res. 1989, 23, 481–486. [Google Scholar] [CrossRef]
- Elser, J.J. The pathway to noxious cyanobacteria blooms in lakes: The food web as the final turn. Freshw. Biol. 1999, 42, 537–543. [Google Scholar] [CrossRef]
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–847. [Google Scholar] [CrossRef]
- Dawson, R.M. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Hamill, K.D. Toxicity in a benthic freshwater cyanobacteria (blue-green algae): First observations in New Zealand. N. Z. J. Mar. Freshw. Res. 2001, 35, 1057–1069. [Google Scholar] [CrossRef]
- Porter, K.G.; Orcutt, J.D. Nutritional adequacy, manageability and toxicity as factors that determine the food quality of green and blue-green algae for Daphnia. In Ecology and Evolution in Zooplankton Communities; Kerfoot, W.C., Ed.; University Press of New England: Hanover, Germany, 1980; pp. 268–281. [Google Scholar]
- Lampert, W. Laboratory studies on zooplankton-cyanobacteria interactions. N. Z. J. Mar. Freshw. Res. 1987, 21, 483–490. [Google Scholar] [CrossRef]
- Sellner, K.G.; Brownlee, D.C.; Bundy, M.H.; Brownlee, S.G.; Braun, K.R. Zooplankton grazing in a Potomac river cyanobacteria bloom. Estuaries 1993, 16, 859–872. [Google Scholar] [CrossRef]
- Sellner, K.G.; Olson, M.M.; Kononen, K. Copepod grazing in a summer cyanobacteria bloom in the Gulf of Finland. Hydrobiologia 1994, 292, 249–254. [Google Scholar] [CrossRef]
- Wilson, A.E.; Hay, M.E. A direct test of cyanobacterial chemical defense: Variable effects of microcystin-treated food on two Daphnia pulicaria clones. Limnol. Oceanogr. 2007, 52, 1467–1479. [Google Scholar] [CrossRef]
- Søndergaard, M.; Liboriussen, L.; Pedersen, A.R.; Jeppesen, E. Lake restoration by fish removal: Long-term effects in 36 Danish lakes. Ecosystems 2008, 11, 1291–1305. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Davidson, T.A.; Liu, Z.; Mazzeo, N.; Trochine, C.; Özkan, K.; Jensen, H.S.; Trolle, D.; et al. Biomanipulation as a restoration tool to combat eutrophication: Recent advances and future challenges. Adv. Ecol. Res. 2012, 47, 411–487. [Google Scholar]
- Hanazato, T.; Yasuno, M. Evaluation of Microcystis as food for zooplankton in a eutrophic lake. Hydrobiologia 1987, 144, 251–259. [Google Scholar] [CrossRef]
- Yu, J.; Li, Y.; Liu, X.; Li, K.; Chen, F.; Gulati, R.; Liu, Z. The fate of cyanobacterial detritus in the food web of Lake Taihu: A mesocosm study using 13C and 15N labeling. Hydrobiologia 2013, 710, 39–46. [Google Scholar] [CrossRef]
- Luo, X.; Liu, Z.; Gulati, R.D. Cyanobacterial carbon supports the growth and reproduction of Daphnia: An experimental study. Hydrobiologia 2015, 743, 211–220. [Google Scholar] [CrossRef]
- De Kluijver, A.; Yu, J.; Houtekamer, M.; Middelburg, J.J.; Liu, Z. Cyanobacteria as a carbon source for zooplankton in eutrophic Lake Taihu, China, measured by 13C labelling and fatty acid biomarkers. Limnol. Oceanogr. 2012, 57, 1245–1254. [Google Scholar] [CrossRef]
- Specziár, A.; Tölg, L.; Bíró, P. Feeding strategy and growth of cyprinids in the littoral zone of Lake Balaton. J. Fish Biol. 1997, 51, 1109–1124. [Google Scholar]
- Qin, B.; Zhu, G.; Gao, G.; Zhang, Y.; Li, W.; Paerl, H.W.; Carmichael, W.W. A drinking water crisis in Lake Taihu, China: Linkage to climatic variability and lake management. Environ. Manag. 2010, 45, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Ma, R.; Xu, X.; Kong, F.; Zhang, S.; Kong, W.; Hao, J.; Shang, L. Two-decade reconstruction of algal blooms in China’s Lake Taihu. Environ. Sci. Technol. 2009, 43, 3522–3528. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Qin, B.; Paerl, H.W.; Brooks, J.D.; Hall, N.S.; Shi, K.; Zhou, Y.; Guo, J.; Li, Z.; Xu, H.; et al. The persistence of cyanobacterial (Microcystis spp.) blooms throughout winter in Lake Taihu, China. Limnol. Oceanogr. 2016, 61, 711–722. [Google Scholar] [CrossRef]
- Yang, G.; Zhong, C.; Pan, H. Comparative studies on seasonal variations of metazooplankton in waters with different eutrophic states in Lake Taihu. Environ. Monit. Assess. 2009, 150, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cai, Y.; Tang, X.; Shao, K.; Qin, B.; Gong, Z. Community structure of macrozoobenthos and the evaluation of water environment in Lake Taihu. J. Lake Sci. 2015, 27, 840–852. (In Chinese) [Google Scholar] [Green Version]
- Cai, Y.; Gong, Z.; Qin, B. Standing crop and spatial distributional pattern of Mollusca in Lake Taihu, 2006–2007. J. Lake Sci. 2009, 21, 713–719. (In Chinese) [Google Scholar]
- Chen, Y.; Fan, C.; Teubner, K.; Dokulil, M. Changes of nutrients and phytoplankton chlorophyll-a in a large shallow lake, Taihu, China: An 8-year investigation. Hydrobiologia 2003, 506, 273–279. [Google Scholar] [CrossRef]
- DeMott, W.R. Food selection by calanoid copepods in response to between-lake variation in food abundance. Freshw. Biol. 1995, 33, 171–180. [Google Scholar] [CrossRef]
- Gophen, M.; Geller, W. Filter mesh size and food particle uptake by Daphnia. Oecologia 1984, 64, 408–412. [Google Scholar] [CrossRef]
- DeMott, W.R. Discrimination between algae and artificial particles by freshwater and marine copepods. Limnol. Oceanogr. 1988, 33, 397–408. [Google Scholar] [CrossRef]
- Repka, S.; van der Vlies, M.; Vijverberg, J. Food quality of detritus derived from the filamentous cyanobacterium Oscillatoria limnetica for Daphnia galeata. J. Plankton Res. 1998, 20, 2199–2205. [Google Scholar] [CrossRef]
- Gulati, R.D.; Bronkhorst, M.; van Donk, E. Feeding in Daphnia galeata on Oscillatoria limnetica and on detritus derived from it. J. Plankton Res. 2001, 23, 705–718. [Google Scholar] [CrossRef]
- Chen, F.; Xie, P. The effects of fresh and decomposed Microcystis aeruginosa on cladocerans from a subtropic Chinese lake. J. Freshw. Ecol. 2003, 18, 97–104. [Google Scholar] [CrossRef]
- Li, K.; Liu, Z.; Li, C.; Li, Y.; Wen, M. Food sources of snail Radix swinhoei in Lake Taihu. J. Lake Sci. 2008, 20, 339–343. (In Chinese) [Google Scholar]
- Zheng, Y.; Wen, M.; Li, K.; Wang, H. Effects of Bellamy sp. on the growth of Vallisneria natans in Lake Taihu. Res. Environ. Sci. 2008, 21, 94–98. (In Chinese) [Google Scholar]
- Levri, E.P.; Lively, C.M. The effects of size, reproductive condition, and parasitism on foraging behaviour in a freshwater snail, Potamopyrgus antipodarum. Anim. Behav. 1996, 51, 891–901. [Google Scholar] [CrossRef]
- Li, K.; Liu, Z.; Hu, Y.; Yang, H. Snail herbivory on submerged macrophytes and nutrient release: Implications for macrophyte management. Ecol. Eng. 2009, 35, 1664–1667. [Google Scholar] [CrossRef]
- Dionisio Pires, L.M.; Bontes, B.M.; Samchyshyna, L.; Jong, J.; Van Donk, E.; Ibelings, B.W. Grazing on microcystin-producing and microcystin-free phytoplankters by different filter-feeders: Implications for lake restoration. Aquat. Sci. 2007, 69, 534–543. [Google Scholar] [CrossRef]
- He, H.; Liu, X.; Liu, X.; Yu, J.; Li, K.; Guan, B.; Jeppesen, E.; Liu, Z. Effects of cyanobacterial blooms on submerged macrophytes alleviated by the native Chinese bivalve Hyriopsis cumingii: A mesocosm experiment study. Ecol. Eng. 2014, 71, 363–367. [Google Scholar] [CrossRef]
- Higgins, S.N.; Vander Zanden, M.J. What a difference a species makes: A meta-analysis of dreissenid mussel impacts on freshwater ecosystems. Ecol. Monogr. 2010, 80, 179–196. [Google Scholar] [CrossRef]
- Kim, B.H.; Lee, J.H.; Hwang, S.J. Inter- and intra-specific differences in filtering activities between two unionids, Anodonta woodiana and Unio douglasiae, in ambient eutrophic lake waters. Ecol. Eng. 2011, 37, 1957–1967. [Google Scholar] [CrossRef]
- Wu, Z.; Qiu, X.; Zhang, X.; Liu, Z.; Tang, Y. Effects of Anodonta woodiana on water quality improvement in restoration of eutrophic shallow lakes. J. Lake Sci. 2018, 30, 1610–1615. (In Chinese) [Google Scholar]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; He, H.; Liu, Z.; Jeppesen, E.; Chen, F.; Zhang, Y. Carbon Transfer from Cyanobacteria to Pelagic and Benthic Consumers in a Subtropical Lake: Evidence from a 13C Labelling Experiment. Water 2019, 11, 1536. https://doi.org/10.3390/w11081536
Yu J, He H, Liu Z, Jeppesen E, Chen F, Zhang Y. Carbon Transfer from Cyanobacteria to Pelagic and Benthic Consumers in a Subtropical Lake: Evidence from a 13C Labelling Experiment. Water. 2019; 11(8):1536. https://doi.org/10.3390/w11081536
Chicago/Turabian StyleYu, Jinlei, Hu He, Zhengwen Liu, Erik Jeppesen, Feizhou Chen, and Yongdong Zhang. 2019. "Carbon Transfer from Cyanobacteria to Pelagic and Benthic Consumers in a Subtropical Lake: Evidence from a 13C Labelling Experiment" Water 11, no. 8: 1536. https://doi.org/10.3390/w11081536