Evapotranspiration from Horizontal Subsurface Flow Constructed Wetlands Planted with Different Perennial Plant Species

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
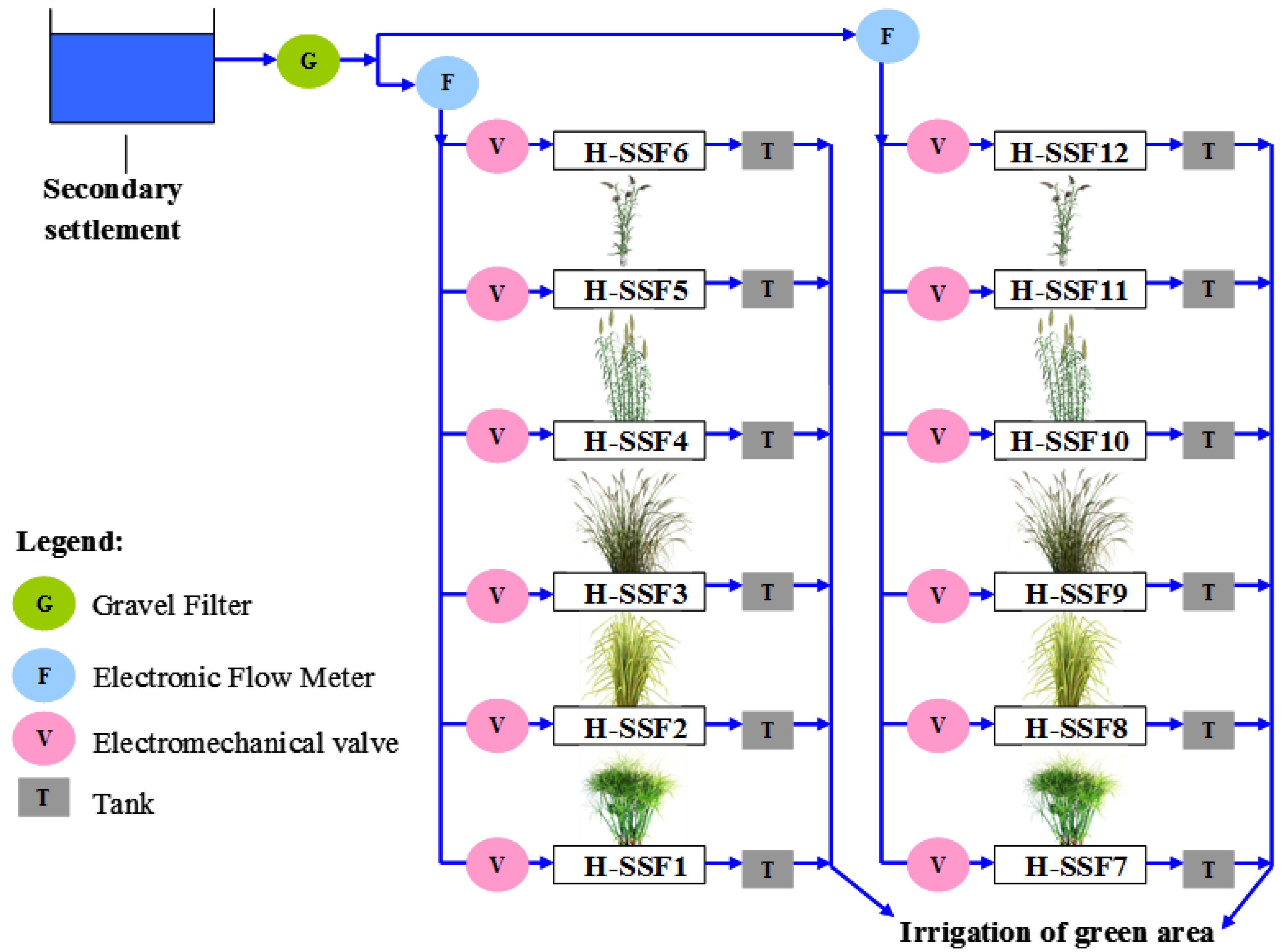
2.1. Experimental Plant Design
2.2. Herbaceous Plants
2.3. Measurement of Bio-Agronomic Characteristics
2.4. Measurement of Evapotranspiration Rates and Crop Coefficients Estimation
2.5. Evaluation of Water Use Efficiency Indices
2.6. Statistical Analysis
3. Results and Discussion
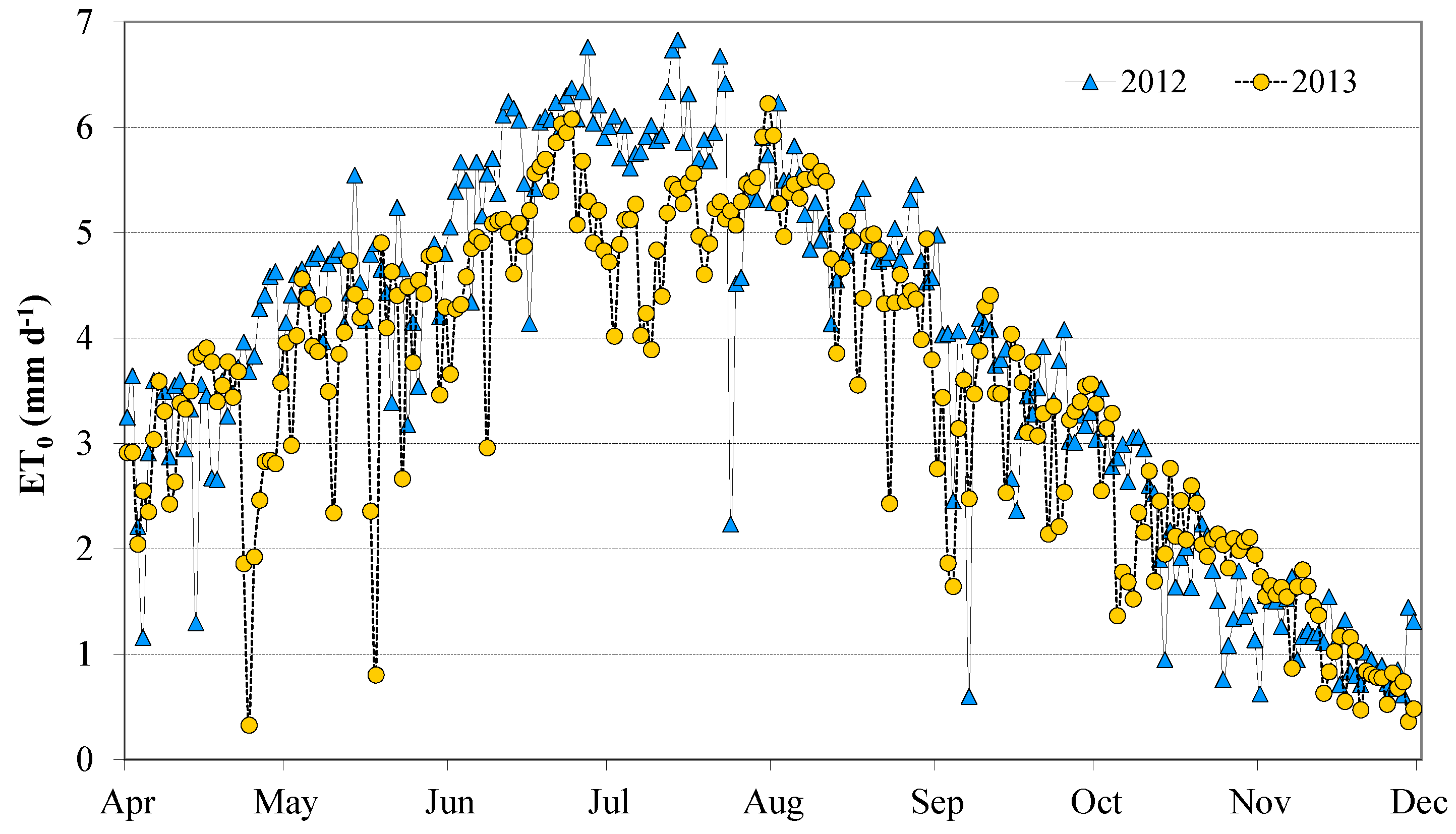
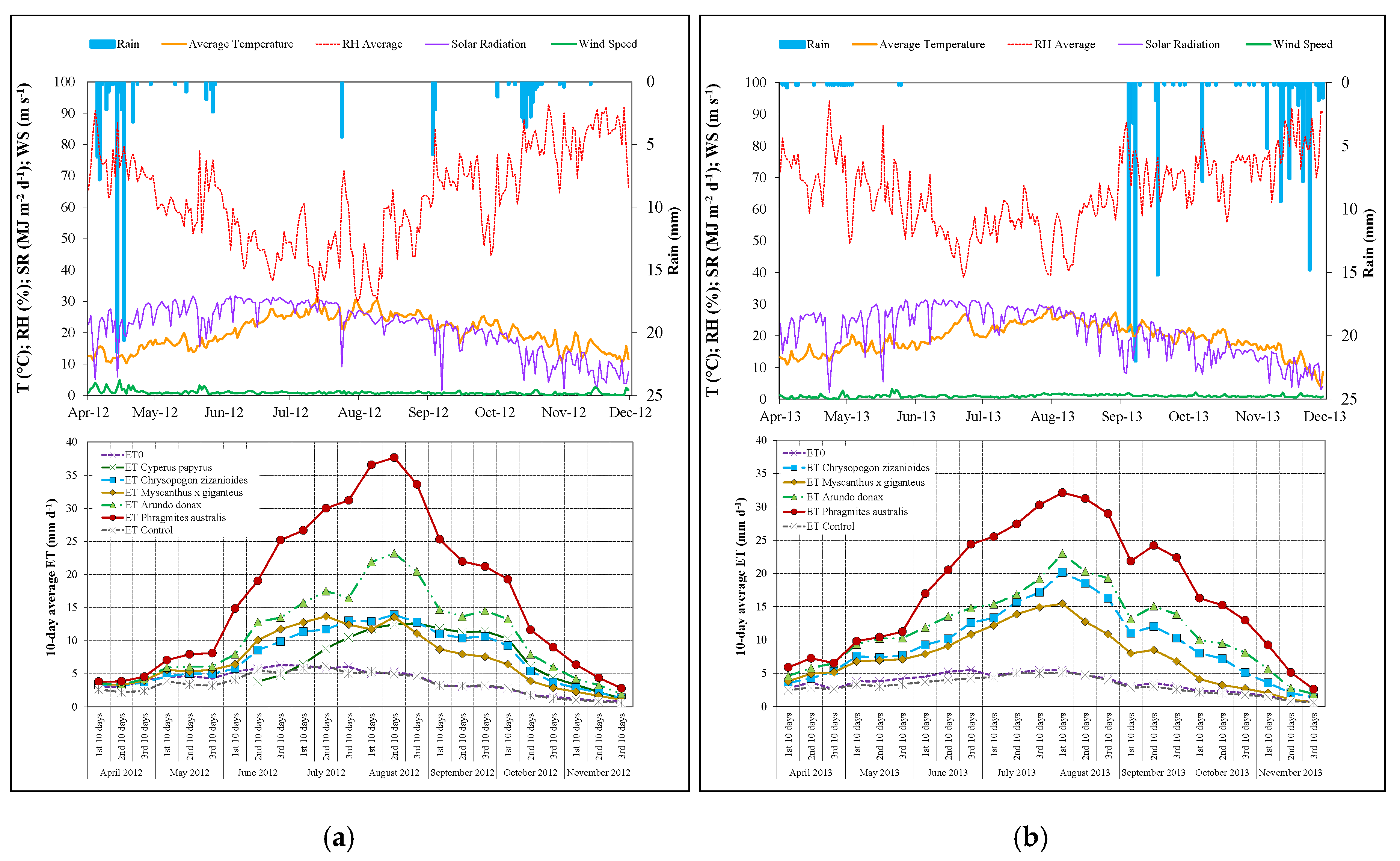
3.1. Meteorological Conditions
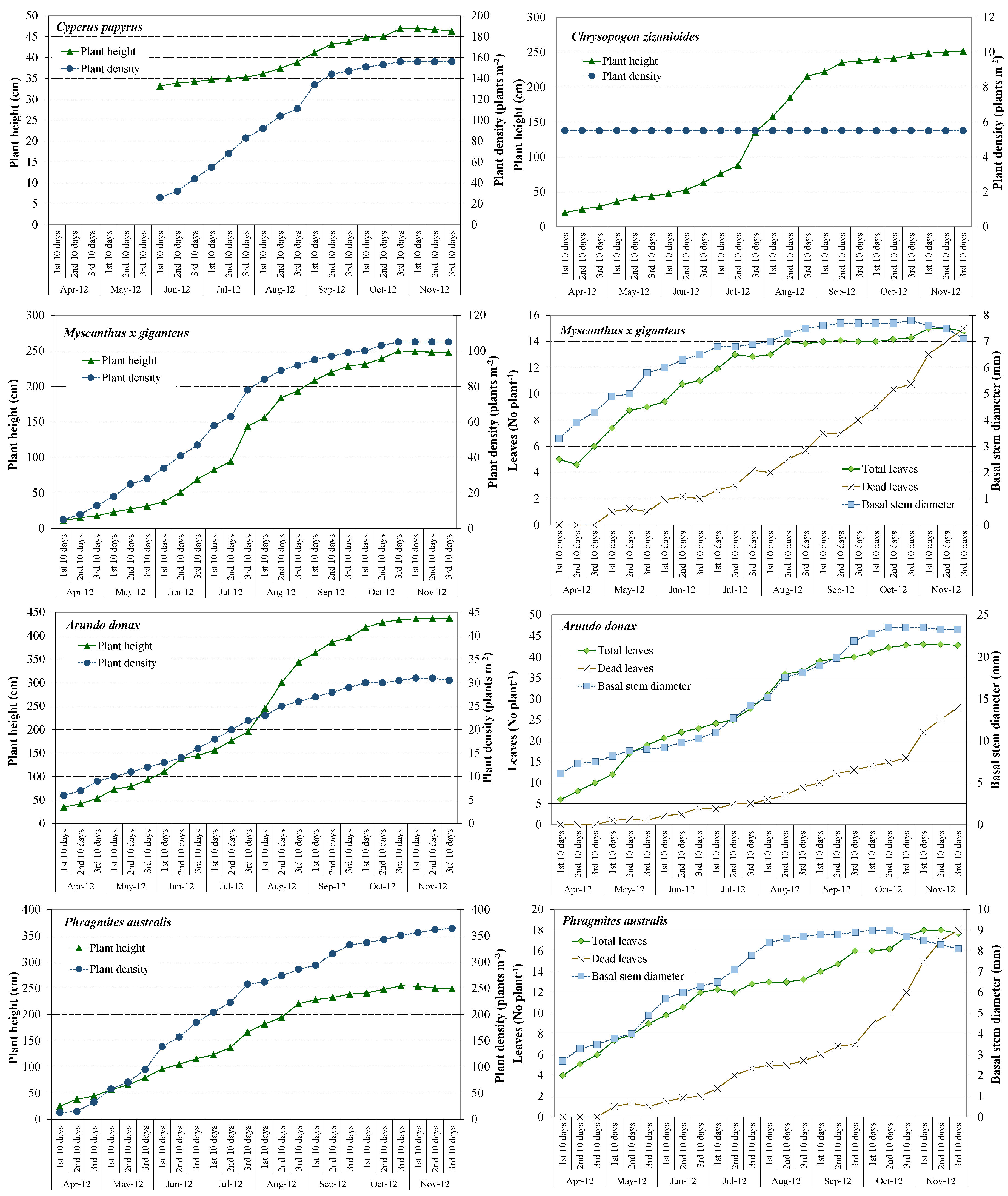
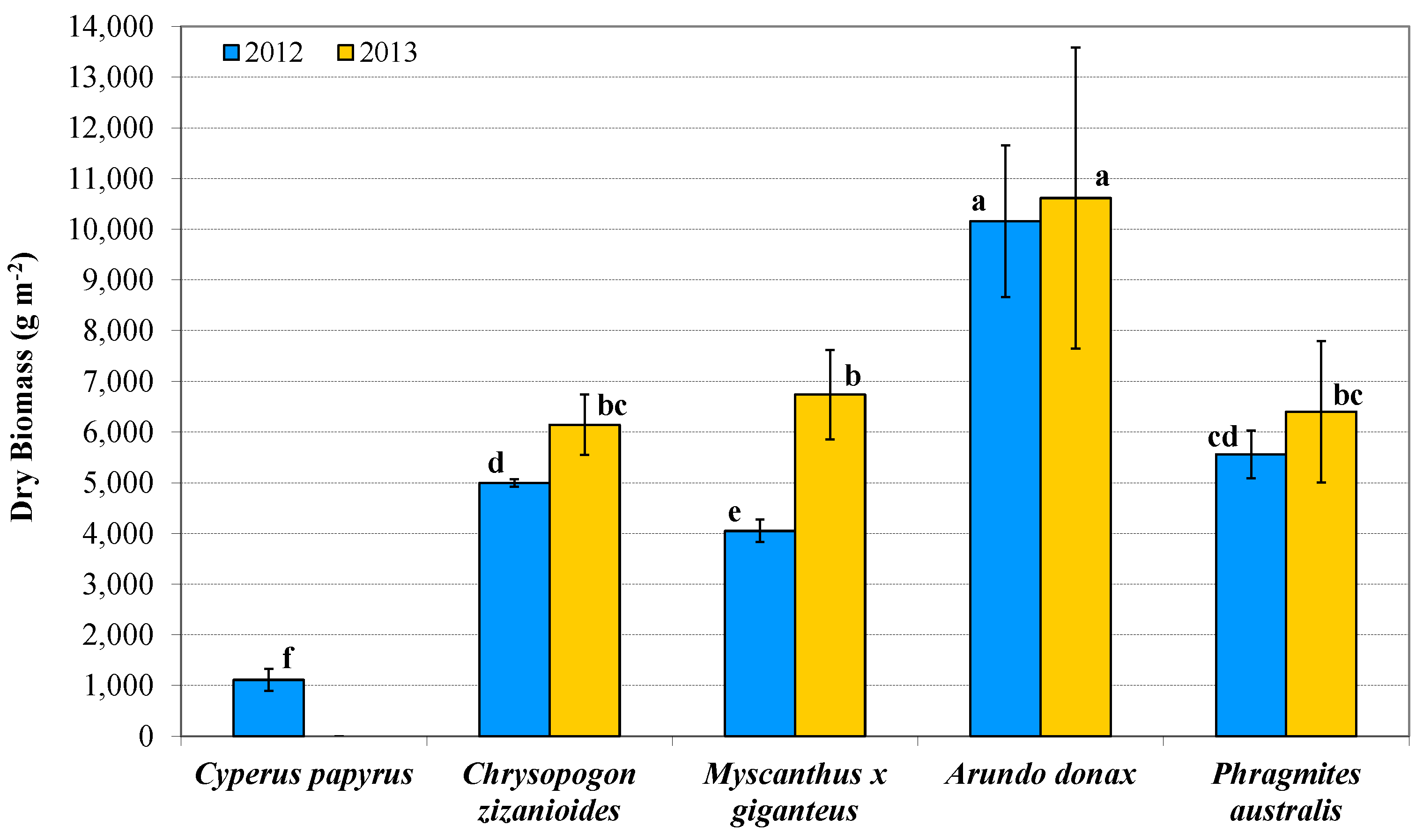
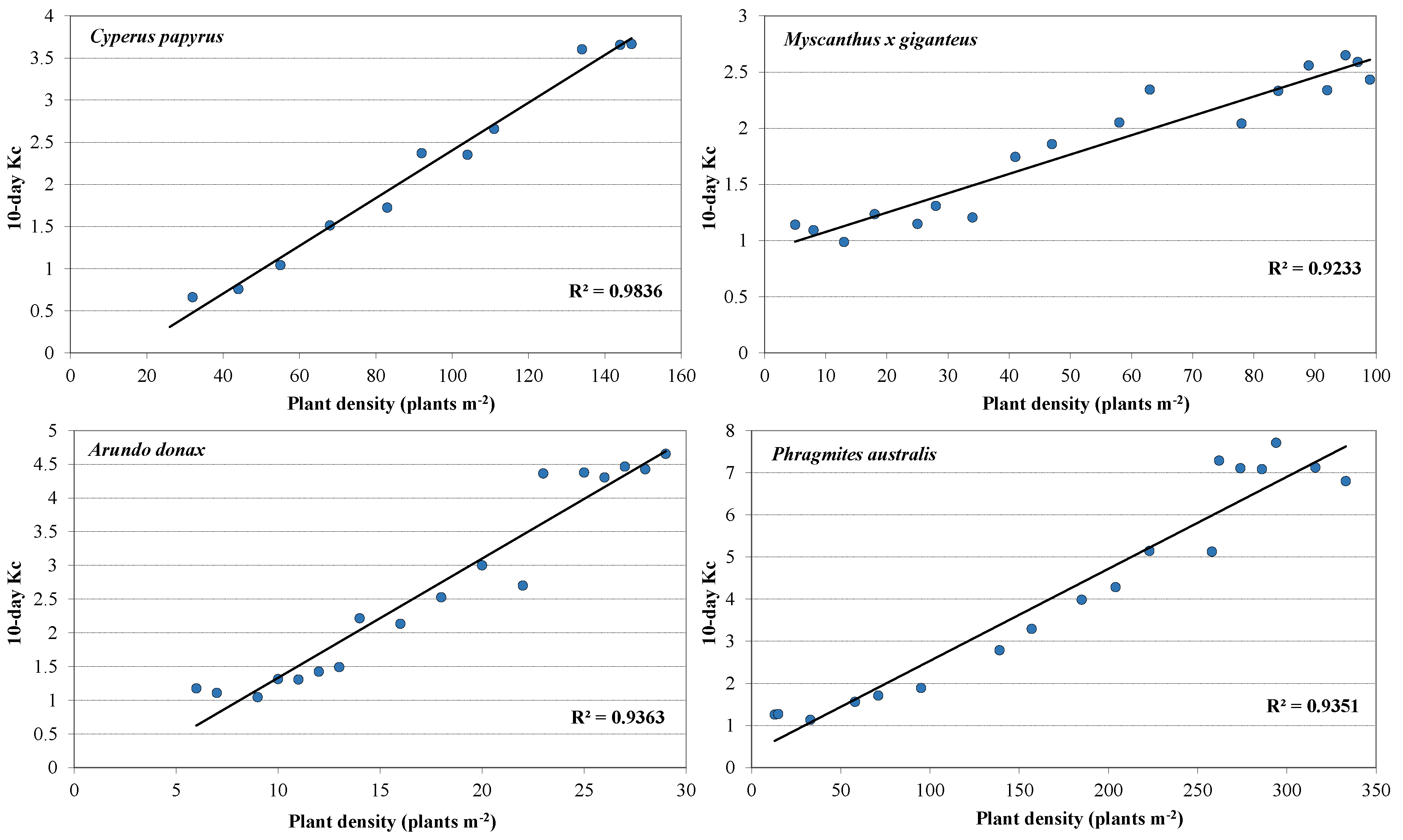
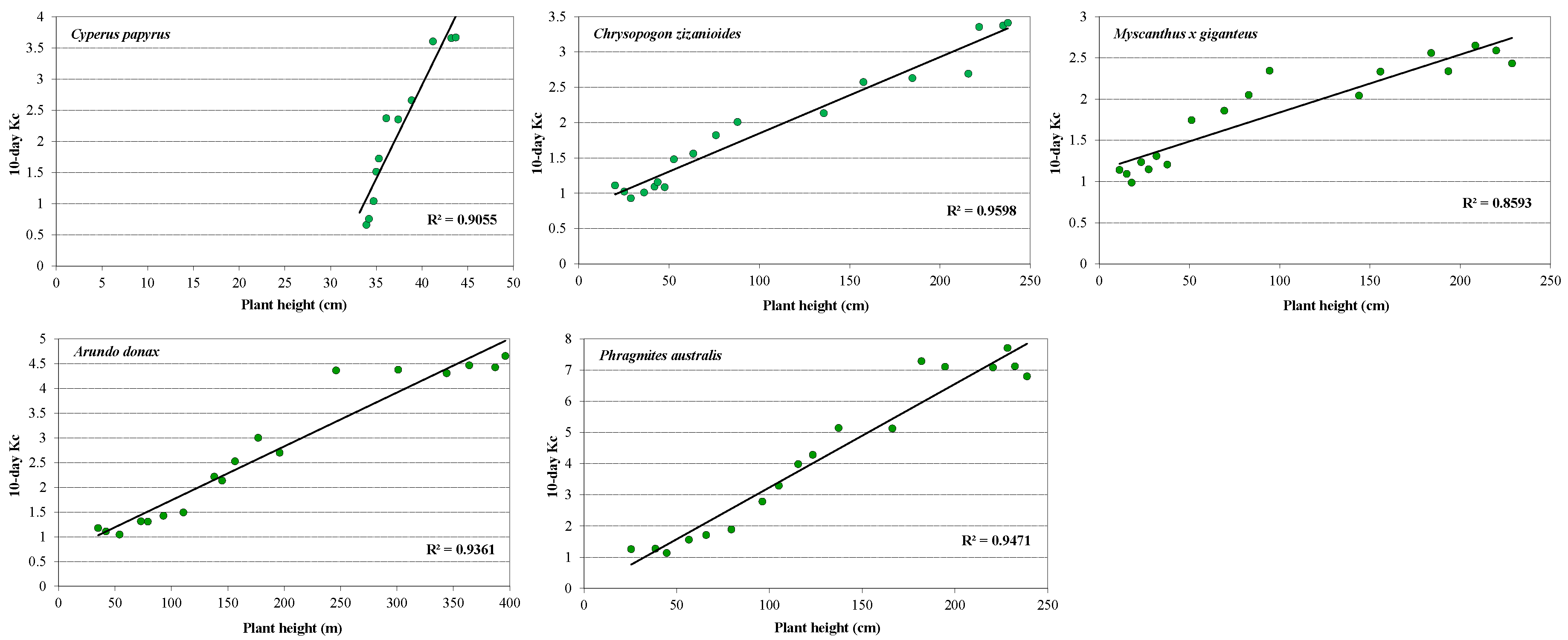
3.2. Plant Development and Biomass Production
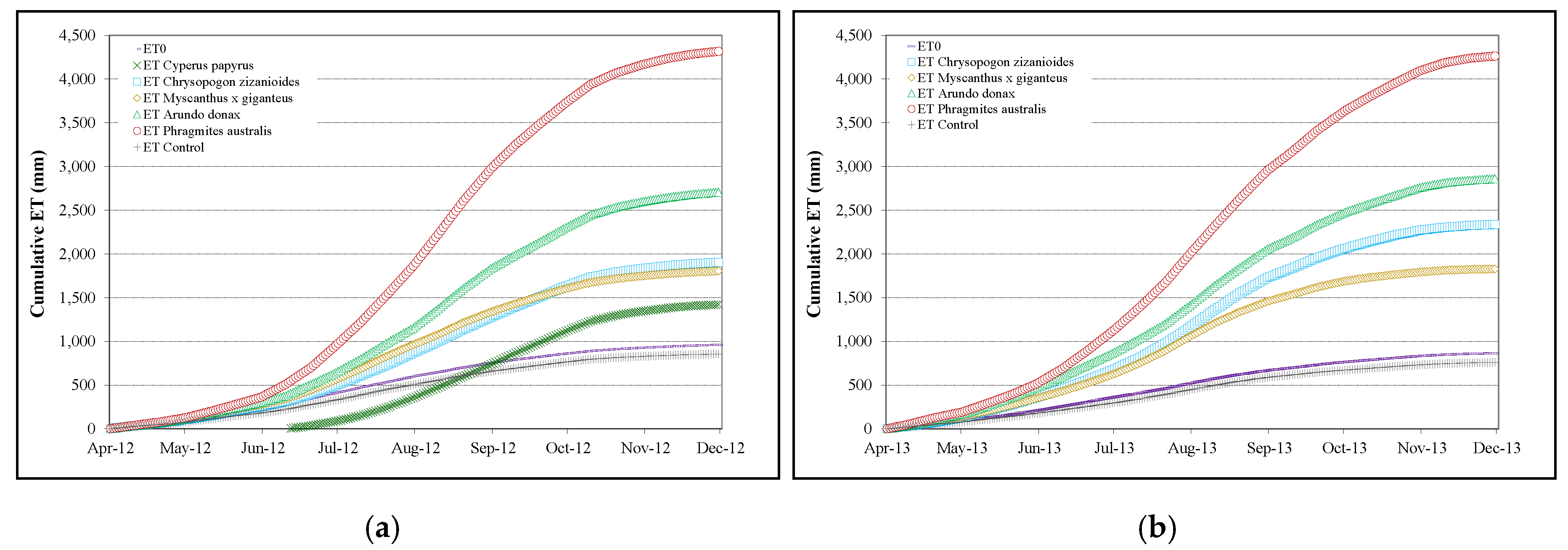
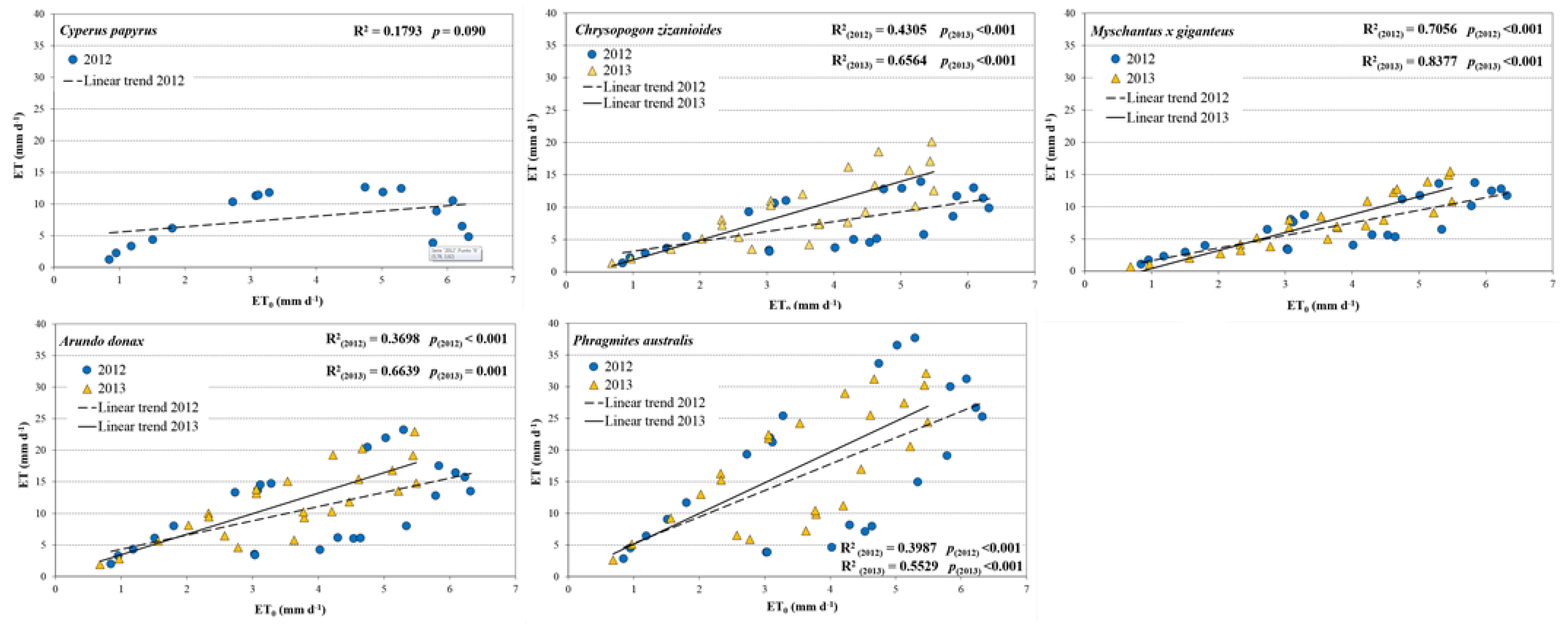
3.3. Evapotranspiration Measurements
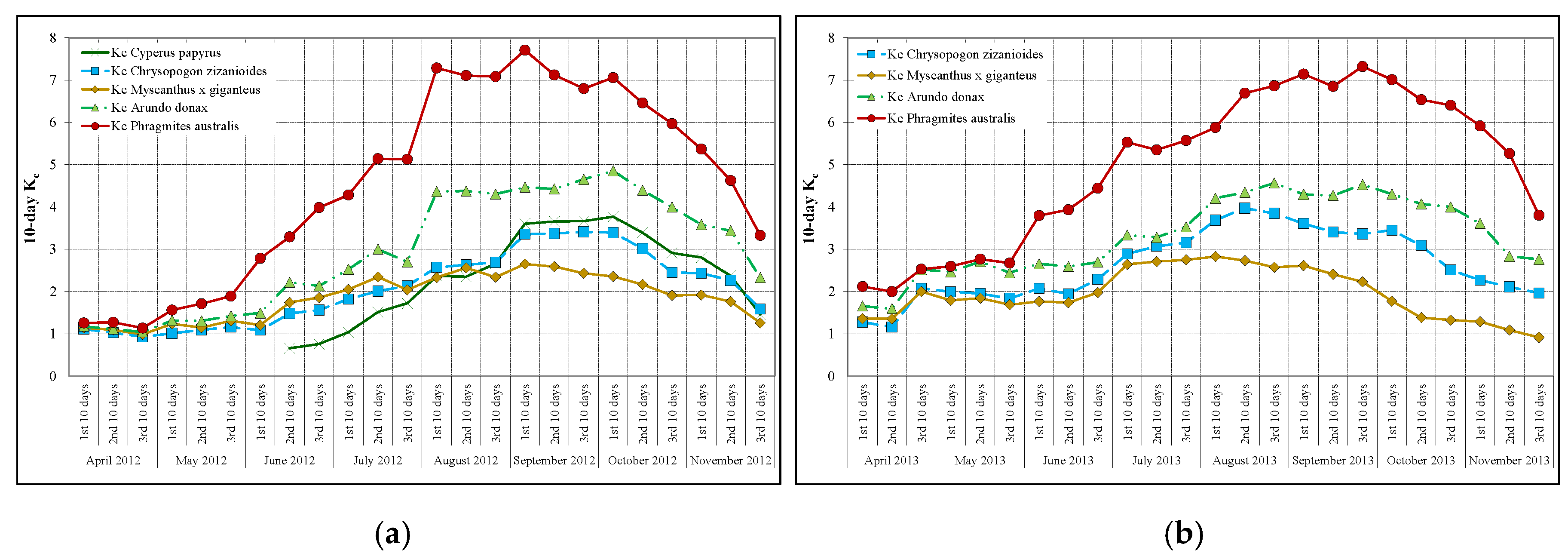
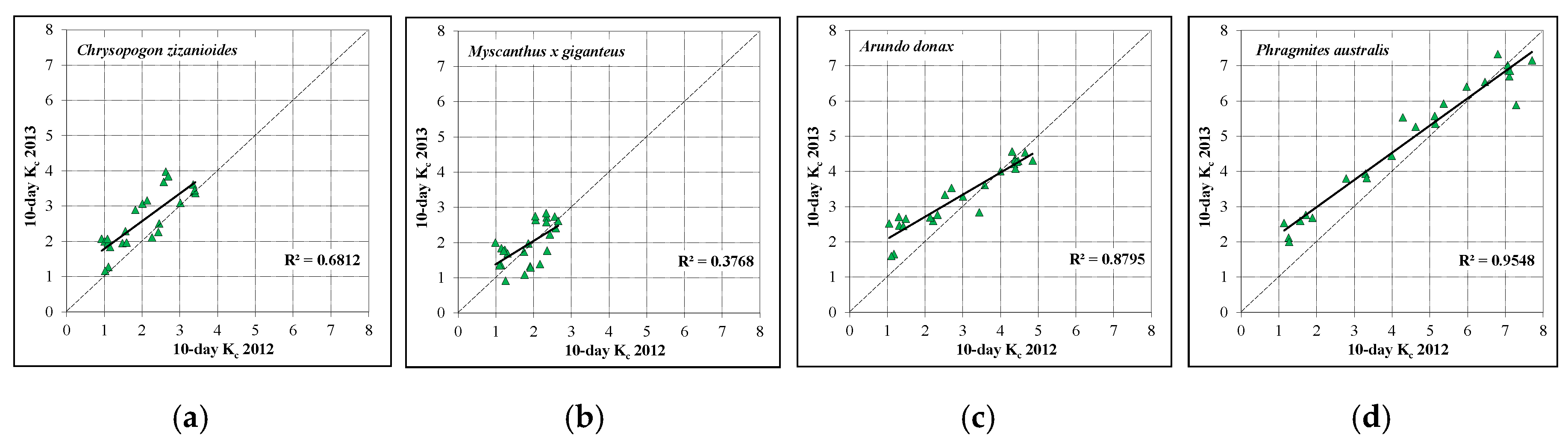
3.4. Crop Coefficients
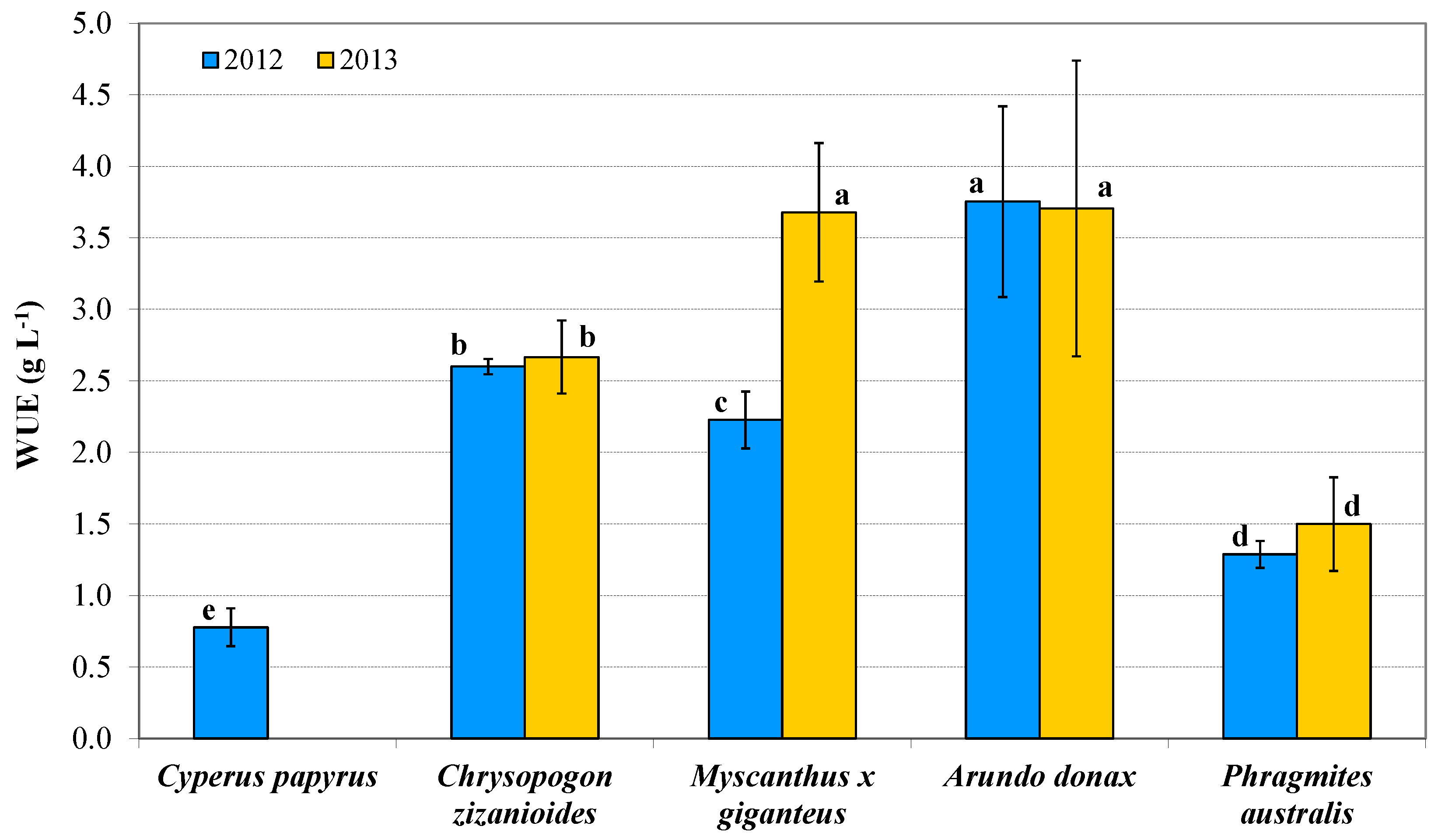
3.5. Water Use Efficiency Indices
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhi, W.; Ji, G. Constructed Wetlands, 1991–2011: A Review of Research Development, Current Trends, and Future Directions. Sci. Total Environ. 2011, 441, 19–27. [Google Scholar] [CrossRef]
- Ventura, D.; Consoli, S.; Barbagallo, S.; Marzo, A.; Vanella, D.; Licciardello, F.; Cirelli, G.L. How to Overcome Barriers for Wastewater Agricultural Reuse in Sicily (Italy)? Water 2019, 11, 335. [Google Scholar] [CrossRef]
- Brisson, J.; Chazarenc, F. Maximizing pollutant removal in constructed wetlands: Should we pay more attention to macrophyte species selection? Sci. Total Environ. 2009, 407, 3923–3930. [Google Scholar] [CrossRef]
- Borin, M.; Milani, M.; Salvato, M.; Toscano, A. Evaluation of Phragmites australis (Cav.) Trin. evapotranspiration in Northern and Southern Italy. Ecol. Eng. 2011, 37, 721–728. [Google Scholar] [CrossRef]
- Milani, M.; Toscano, A. Evapotranspiration from pilot-scale constructed wetlands planted with Phragmites australis in a Mediterranean environment. J. Environ. Sci. Health Part A 2013, 48, 568–580. [Google Scholar] [CrossRef] [PubMed]
- El Hamouri, B.; Nazih, J.; Lahjouj, J. Subsurface-horizontal flow constructed wetland for sewage treatment under Moroccan climate conditions. Desalination 2007, 215, 153–158. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Wallace, S.D. Treatment Wetlands, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009; 1016p. [Google Scholar]
- Fiener, P.; Auerswald, K.; Weigand, S. Managing erosion and water quality in agricultural water-sheds by small detention ponds. Agric. Ecosyst. Environ. 2005, 110, 132–142. [Google Scholar] [CrossRef]
- Xu, S.; Jaffè, P.R. Effects of plants on the removal of hexavalent chromium in wetland sediments. J. Environ. Qual. 2006, 35, 334–341. [Google Scholar] [CrossRef]
- Brix, H. Sludge Dewatering and Mineralization in Sludge Treatment Reed Beds. Water 2017, 9, 160. [Google Scholar] [CrossRef]
- Toscano, A.; Marzo, A.; Milani, M.; Cirelli, G.L.; Barbagallo, S. Comparison of removal efficiencies in Mediterranean pilot constructed wetlands vegetated with different plant species. Ecol. Eng. 2015, 75, 155–160. [Google Scholar] [CrossRef]
- Tuttolomondo, T.; Licata, M.; Leto, C.; Leone, R.; La Bella, S. Effect of plant species on water balance in a pilot-scale horizontal subsurface flow constructed wetland planted with Arundo donax L. and Cyperus alternifolius L.—Two-year tests in a Mediterranean environment in the West of Sicily (Italy). Ecol. Eng. 2015, 75, 79–92. [Google Scholar] [CrossRef]
- Toscano, A.; Hellio, C.; Marzo, A.; Milani, M.; Lebret, K.; Cirelli, G.L.; Langergraber, G. Removal efficiency of a constructed wetland combined with ultrasound and UV devices for wastewater reuse in agriculture. Environ. Technol. 2013, 34, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, S.; Cirelli, G.L.; Consoli, S.; Licciardello, F.; Marzo, A.; Toscano, A. Analysis of treated wastewater reuse potential for irrigation in Sicily. Water Sci. Technol. 2012, 65, 2024–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consoli, S.; Milani, M.; Cirelli, G.L.; Barbagallo, S.; Marzo, A.; Vanella, D.; Toscano, A. Energy and water balance of a treatment wetland under mediterranean climatic conditions. Ecol. Eng. 2018, 116, 52–60. [Google Scholar] [CrossRef]
- Headley, T.R.; Davison, L.; Huett, D.O.; Müller, R. Evapotranspiration from subsurface horizontal flow wetlands planted with Phragmites australis in sub-tropical Australia. Water Res. 2012, 46, 345–354. [Google Scholar] [CrossRef]
- Anda, A.; Teixeira da Silva, J.A.; Soos, G. Evapotranspiration and crop coefficient of common reed at the surroundings of Lake Balaton, Hungary. Aquat. Bot. 2014, 116, 53–59. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, G. Measurement and modelling of evapotranspiration over a reed (Phragmites australis) marsh in Northeast China. J. Hydrol. 2009, 372, 41–47. [Google Scholar] [CrossRef]
- Borin, M.; Salvato, M. Effects of five macrophytes on nitrogen remediation and mass balance in wetland mesocosms. Ecol. Eng. 2012, 46, 34–42. [Google Scholar] [CrossRef]
- Calheiros, C.S.C.; Rangel, A.O.S.S.; Castro, P.M.L. Constructed wetland systems vegetated with different plants applied to the treatment of tannery wastewater. Water Res. 2007, 41, 1790–1798. [Google Scholar] [CrossRef]
- Pauliukonis, N.; Schneider, R. Temporal patterns in evapotranspiration from lysimeters with three common wetland plant species in the eastern United States. Aquat. Bot. 2001, 71, 35–46. [Google Scholar] [CrossRef]
- Papaevangelou, V.A.; Gikas, G.D.; Tsihrintzis, V.A. Evaluation of evapotranspiration in small on-site HSF constructed wetlands. J. Environ. Sci. Health A. 2012, 47, 766–785. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, S.; Cirelli, G.L.; Marzo, A.; Milani, M.; Toscano, A. Effect of different plant species in pilot constructed wetlands for wastewater reuse in agricultural. J. Agric. Eng. 2013, 44, 796–802. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements; FAO: Rome, Italy, 1998; 300p. [Google Scholar]
- Pedescoll, A.; Sidrach-Cardona, R.; Sánchez, J.C.; Bécares, E. Evapotranspiration affecting redox conditions in horizontal constructed wetlands under Mediterranean climate: Influence of plant species. Ecol. Eng. 2013, 58, 335–343. [Google Scholar] [CrossRef]
- Drexler, J.Z.; Anderson, F.E.; Snyder, R.L. Evapotranspiration rates and crop coefficients for a restored marsh in the Sacramento-San Joaquin Delta, California, USA. Hydrol. Process. 2008, 22, 725–735. [Google Scholar] [CrossRef]
- Barbera, A.C.; Cirelli, G.L.; Di Silvestro, I.; Pacifici, P.; Castiglione, V.; Toscano, A.; Milani, M. Growth and biomass production of different plant species in two different constructed wetland systems in Sicily. Desalination 2009, 246, 129–136. [Google Scholar] [CrossRef]
- Means, M.M.; Ahn, C.; Noe, G.B. Planting richness affects the recovery of vegetation and soil processes in constructed wetlands following disturbance. Sci. Total Environ. 2017, 579, 1366–1378. [Google Scholar] [CrossRef] [Green Version]
- Vymazal, J.; Kröpfelová, L. Growth of Phragmites australis and Phalaris arundinacea in constructed wetlands for wastewater treatment in the Czech Republic. Ecol. Eng. 2005, 25, 606–621. [Google Scholar] [CrossRef]
- Zygas, A.; Eheart, J.; Cai, X. Simulation and Optimization of a Constructed Wetland for Biomass Production and Nitrate Removal. J. Water Resour. Plan. Manag. 2014, 140, 1–10. [Google Scholar] [CrossRef]
- APHA, AWWA, AEF. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA), American WaterWorks Association (AWWA), and American Environment Federation (AEF): Washington, DC, USA, 2005. [Google Scholar]
- Rossa, B.; Tuffers, A.V.; Naidoo, G.; Von Willert, D.J. Arundo donax L. (Poaceae) a C3 species with unusually high photosynthetic capacity. Bot. Acta 1998, 111, 216–221. [Google Scholar] [CrossRef]
- Lewandowski, I.; Scurlock, J.M.O.; Lindvall, E.; Christou, M. The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass Bioenergy 2003, 25, 335–361. [Google Scholar] [CrossRef]
- Czako, M.; Marton, L. Subtropical and Tropical Reeds for Biomass (Chapter 16). In Energy Crops; The Royal Society of Chemistry: London, UK, 2010; pp. 322–340. [Google Scholar]
- Dudley, T.L. Arundo donax L. In Invasive Plants of California’s Wildlands; Bossard, C.C., Randall, J.M., Hoshovsky, M.C., Eds.; University of California Press: Berkeley, CA, USA, 2000; pp. 53–58. [Google Scholar]
- Idris, S.M.; Jones, P.L.; Salzman, S.A.; Croatto, G.; Allison, G. Evaluation of the giant reed (Arundo donax) in horizontal subsurface flow wetlands for the treatment of recirculating aquaculture system effluent. Environ. Sci. Pollut. Res. 2012, 19, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Greef, J.M.; Deuter, M. Syntaxonomy of Miscanthus x giganteus GREEF et DEU. Angew. Bot. 1993, 67, 87–90. [Google Scholar]
- Hodkinson, T.R.; Renvoize, S.A.; Chase, M.W. Systematics in Miscanthus. Asp. Appl. Biol. 1997, 49, 189–198. [Google Scholar]
- Anderson, E.; Arundale, R.; Maughan, M.; Oladeinde, A.; Wycislo, A.; Voigt, T. Growth and agronomy of Miscanthus x giganteus for biomass production. Biofuels 2011, 2, 167–183. [Google Scholar] [CrossRef]
- Stander, W. Determination of the Highest Biomass Producing Plant Genera (C4 Grasses) of the World for Temperate Climates; Assessment Study for the Commission of the European Communities: Munich, Germany, 1989; 74p. [Google Scholar]
- Beale, C.V.; Bint, D.A.; Long, S.P. Leaf photosynthesis in the C4-grass Miscanthus x giganteus, growing in the cool temperate climate of southern England. J. Exp. Bot. 1996, 47, 267–273. [Google Scholar] [CrossRef]
- Vymazal, J. Plants used in constructed wetlands with horizontal subsurface flow: A review. Hydrobiologia 2011, 674, 133–156. [Google Scholar] [CrossRef]
- Garcia, J.; Aguirre, P.; Mueriego, R.; Hung, Y.; Ortiz, L.; Bayona, J.M. Initial contamination removal performance factor in horizontal flow reed beds used for treating urban wastewater. Water Res. 2004, 38, 1669–1678. [Google Scholar] [CrossRef]
- Worman, A.; Kronnas, V. Effect of pond shape and vegetation heterogeneity on flow and treatment performance of constructed wetlands. J. Hydrol. 2005, 301, 123–138. [Google Scholar] [CrossRef]
- Maltais-Landry, G.; Maranger, R.; Brisson, J.; Chazarenc, F. Nitrogen transformations and retention in planted and artificially aerated constructed wetlands. Water Res. 2009, 43, 535–545. [Google Scholar] [CrossRef]
- Xia, H.P. Ecological rehabilitation and phytoremediation with four grasses in oil shale mined land. Chemosphere 2004, 54, 345–353. [Google Scholar] [CrossRef]
- Jones, M.B. The photosynthetic characteristics of papyrus in a tropical swamp. Oecologia 1987, 71, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Kuekenthal, G. Cyperaceae–Scirpoideae-Cyperae. In Das Pflanzenreich IV; 20 (Heft 101); Engler, A., Ed.; Wihelm Engleman: Leipzig, Germany, 1935; 671p. [Google Scholar]
- De Filipps, R.A. Cyperus. In Flora Europea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; pp. 284–288. [Google Scholar]
- Gordon-Gray, K.D. Cyperaceae in Natal; National Botanical Institute: Pritoria, South Africa, 1995; 218p. [Google Scholar]
- Mabberley, D.J. The Plant Book. A Portable Dictionary of the Vascular Plants, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997; 865p. [Google Scholar]
- Wunderlin, R.P. Guide to the Vascular Plants of Florida; University Press of Florida: Gainesville, FL, USA, 1998; 806p. [Google Scholar]
- Snyder, R.L.; Eching, S. Daily ETos, ETrs and HS ETo Calculator Using the ASCE-EWRI Method.xls; University of California, Davis: Davis, CA, USA, 2007; Available online: http://biomet.ucdavis.edu/Evapotranspiration/PMdayXLS/PMday.xls (accessed on 4 September 2019).
- Snyder, R.L.; Eching, S. Penman-Monteith Daily (24-h) Reference Evapotranspiration Equations for Estimating ETo, ETr and HS ETo with Daily Data; University of California, Davis: Davis, CA, USA, 2009; Available online: http://biomet.ucdavis.edu/Evapotranspiration/PMdayXLS/PMdayDoc.pdf (accessed on 4 September 2019).
- ASCE-EWRI. The ASCE Standardized Reference Evapotranspiration Equation. In Environmental and Water Resources Institute (EWRI) of the American Society of Civil Engineers (ASCE) Standardization of Reference Evapotranspiration Task Committee Final Report; Allen, R.G., Walter, I.A., Elliott, R., Howell, T.A., Itenfisu, D., Jensen, M.E., Eds.; ASCE-EWRI: Reston, VA, USA, 2005; 213p. [Google Scholar]
- Sinclair, T.R.; Tanner, C.B.; Bennett, J.M. Water use efficiency in crop production. Bioscience 1984, 34, 40–60. [Google Scholar] [CrossRef]
- Borin, M.; Barbera, A.C.; Milani, M.; Molari, G.; Zimbone, S.M.; Toscano, A. Biomass production and N balance of giant reed (Arundo donax L.) under high water and N input in Mediterranean environments. Eur. J. Agron. 2013, 51, 117–119. [Google Scholar] [CrossRef]
- Kuusemets, V.; Lõhmus, K. Nitrogen and Phosphorus Accumulation and Biomass Production by Scirpus sylvaticus and Phragmites australis in a Horizontal Subsurface Flow Constructed Wetland. J. Environ. Sci. Health Part A 2005, 40, 1167–1175. [Google Scholar] [CrossRef]
- Mueller, L.; Behrendt, A.; Schalitz, G.; Schindler, U. Above ground biomass and water use efficiency of crops at shallow water tables in a temperate climate. Agric. Water Manag. 2005, 25, 117–136. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H. Monthly variations of trace elements accumulation and distribution in above- and below-ground biomass of Phragmites australis (Cav.) Trin. Ex Steudel in Lake Burullus (Egypt): A biomonitoring application. Ecol. Eng. 2014, 73, 17–25. [Google Scholar] [CrossRef]
- Zub, H.W.; Brancourt-Hulmel, M. Agronomic and physiological performances of different species of Miscanthus, a major energy crop. A Review. Agron. Sustain. Dev. 2010, 30, 201–214. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, W.; Zeng, Z.; Li, H.; Yang, X.; He, Z.; Gu, B.; Rafiq, M.T.; Peng, H. Nutrient removal efficiency and biomass production of different bioenergy plants in hypereutrophic water. Biomass Bioenergy 2012, 42, 212–218. [Google Scholar] [CrossRef]
- Truong, P.; Smeal, C. Research, Development and Implementation of Vetiver System for Wastewater Treatment: GELITA Australia; Technical Bulletin No. 2003/3; Pacific Rim Vetiver Network. Office of the Royal Development Projects Board: Bangkok, Thailand, 2003. [Google Scholar]
- Perbangkhem, T.; Polprasert, C. Biomass production of papyrus (Cyperus papyrus) in constructed wetland treating low-strength domestic wastewater. Bioresour. Technol. 2010, 101, 833–835. [Google Scholar] [CrossRef]
- Mnaya, B.; Asaeda, T.; Kiwango, Y.; Ayubu, E. Primary production in papyrus (Cyperus papyrus L.) of Rubondo Island, Lake Victoria, Tanzania. Wetl. Ecol. Manag. 2007, 15, 269–275. [Google Scholar] [CrossRef]
- Rozkošný, M.; Šálek, J.; Šálek, J. Water balance of the constructed wetlands – a study of the macrophyte´s evapotranspiration. In Proceedings of the 10th International Conference Wetland Systems for Water Pollution Control, Lisbon, Portugal, 23–29 September 2006. [Google Scholar]
- Herbst, M.; Kappen, L. The ratio of transpiration versus evaporation in a reed belt as influenced by weather conditions. Aquat. Bot. 1999, 63, 113–125. [Google Scholar] [CrossRef]
- Fermor, P.M.; Hedges, P.D.; Gilbert, J.C.; Gowing, D.J.G. Reedbed evapotranspiration rates in England. Hydrol. Process. 2001, 15, 621–631. [Google Scholar] [CrossRef]
- Chazarenc, F.; Merlin, G.; Gonthier, Y. Hydrodynamics of horizontal subsurface flow constructed wetlands. Ecol. Eng. 2003, 21, 165–173. [Google Scholar] [CrossRef]
- Triana, F.; Nassi o Di Nasso, N.; Ragaglini, G.; Roncucci, N.; Bonari, E. Evapotranspiration, crop coefficient and water use efficiency of giant reed (Arundo donax L.) and miscanthus (Miscanthus x giganteus Geef et Deu.) in a Mediterranean environment. GBC Bioenergy 2014, 7, 811–819. [Google Scholar]
- Kyambadde, J.; Kansiime, F.; Dalhammar, G. Nitrogen and phosphorus removal in substrate-free pilot constructed wetlands with horizontal surface flow in Uganda. Water Air Soil Pollut. 2005, 165, 37–59. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Howell, T.A.; Jensen, M.E. Evapotranspiration information reporting: I. Factors governing measurement accuracy. Agric. Water Manag. 2011, 98, 899–920. [Google Scholar] [CrossRef] [Green Version]
- Peacock, C.E.; Hess, T.M. Estimating evapotranspiration from a reed bed using the Bowen ratio energy balance method. Hydrol. Process. 2004, 18, 247–260. [Google Scholar] [CrossRef]
- Stanhill, G. Water use efficiency. Adv. Agron. 1986, 39, 53–85. [Google Scholar]
- Berndes, G. Bioenergy and water—The implications of large-scale bioenergy production for water use and supply. Glob. Environ. Chang. 2002, 12, 253–271. [Google Scholar] [CrossRef]
- Barbagallo, S.; Barbera, A.C.; Cirelli, G.L.; Milani, M.; Toscano, A. Reuse of constructed wetland effluents for irrigation of energy crops. Water Sci. Technol. 2014, 70, 1465–1472. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Patanè, C.; Sanzone, E.; Copani, V.; Foti, S. Effects of soil water content and nitrogen supply on the productivity of Miscanthus x giganteus Greef et Deu. in a Mediterranean environment. Ind. Crops Prod. 2007, 25, 75–88. [Google Scholar] [CrossRef]
- Mantineo, M.; D’Agosta, G.M.; Copani, V.; Patanè, C.; Cosentino, S.L. Biomass yield and energy balance of three perennial crops for energy use in the semi-arid Mediterranean environment. Field Crops Res. 2009, 114, 204–213. [Google Scholar] [CrossRef]
- Christou, M.; Mardikis, M.; Alexopoulou, E.; Cosentino, S.L.; Copani, V.; Sanzone, E. Environmental studies on Arundo donax. In Proceedings of the 8th International Conference on Environmental Science and Technology, Lemnos Island, Greece, 8–10 September 2003; Curran Associates Inc.: Red Hook, NY, USA, 2003; pp. 102–110. [Google Scholar]
- Morrison, E.H.J.; Harper, D.M. Ecohydrological principles to underpin the restoration of Cyperus papyrus at Lake Naivasha, Kenya. Ecohydrol. Hydrobiol. 2009, 9, 83–97. [Google Scholar] [CrossRef]
- Aiello, R.; Cirelli, G.L.; Consoli, S.; Licciardello, F.; Toscano, A. Risk assessment of treated municipal wastewater reuse in Sicily. Water Sci. Technol. 2013, 67, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Ayers, R.S.; Westcot, D.W. Water Quality for Agriculture; FAO Irrigation and Drainage, Paper 29; Food and Agriculture Organization: Rome, Italy, 1985. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Year | |||
|---|---|---|---|---|---|
| 2012 | 2013 | ||||
| Mean | ±SD | Mean | ±SD | ||
| TSS | mg L−1 | 78.0 | (±61.4) | 35.5 | (±16.7) |
| BOD5 | mg L−1 | not detected | 30.8 | (±3.9) | |
| COD | mg L−1 | 67.9 | (±40.4) | 50.6 | (±2.6) |
| NH4-N | mg L−1 | 7.1 | (±4.5) | 11.4 | (±4.8) |
| NO2-N | mg L−1 | 0.3 | (±0.2) | 0.3 | (±0.1) |
| NO3-N | mg L−1 | 6.7 | (±4.7) | 1.0 | (±0.4) |
| Norg | mg L−1 | 1.4 | (±0.7) | 2.4 | (±0.9) |
| TN | mg L−1 | 15.6 | (±7.5) | 15.1 | (±4.6) |
| PO4-P | mg L−1 | 3.3 | (±1.3) | 7.5 | (±0.9) |
| Year | Month | Cumulative Rain | Absolute Minimum Air Temperature | Absolute Maximum Air Temperature | Average Air Temperature | Average Wind Speed | Average Relative Humidity | Average Solar Radiation |
|---|---|---|---|---|---|---|---|---|
| mm | °C | °C | °C | ms−1 | % | MJm−2d−1 | ||
| 2012 | April | 66.0 | 1.5 | 27.1 | 13.3 | 1.8 | 73.9 | 22.2 |
| May | 5.6 | 3.9 | 29.5 | 16.8 | 1.0 | 61.9 | 26.7 | |
| June | 0.0 | 8.2 | 37.5 | 23.1 | 0.9 | 48.1 | 29.9 | |
| July | 4.4 | 13.4 | 43.4 | 26.3 | 0.8 | 47.5 | 27.6 | |
| August | 0.0 | 13.9 | 42.2 | 26.1 | 0.8 | 49.2 | 24.2 | |
| September | 8.0 | 11.7 | 35.6 | 22.0 | 0.7 | 66.0 | 19.6 | |
| October | 18.2 | 3.3 | 34.5 | 19.0 | 0.6 | 75.3 | 13.5 | |
| November | 0.6 | 3.4 | 25.6 | 14.4 | 0.7 | 83.5 | 8.4 | |
| Average Season | 102.8 * | 12.5 | 28.1 | 20.1 | 0.9 | 63.1 | 5253.2 * | |
| 2013 | April | 3.0 | 3.5 | 25.3 | 13.7 | 0.6 | 73.9 | 21.3 |
| May | 1.0 | 4.4 | 30.5 | 17.0 | 1.0 | 64.1 | 24.8 | |
| June | 0.0 | 7.8 | 37.3 | 20.9 | 0.9 | 52.6 | 29.7 | |
| July | 0.0 | 10.2 | 39.9 | 23.9 | 0.9 | 54.9 | 27.3 | |
| August | 0.0 | 14.4 | 37.4 | 24.8 | 1.4 | 57.7 | 24.1 | |
| September | 62.4 | 8.8 | 34.3 | 21.1 | 1.1 | 71.1 | 18.8 | |
| October | 9.2 | 6.4 | 33.6 | 19.2 | 0.9 | 71.9 | 14.5 | |
| November | 61.4 | −1.6 | 26.7 | 12.4 | 0.9 | 79.2 | 9.6 | |
| Average Season | 137.0 * | 11.8 | 27.0 | 19.2 | 1.0 | 65.6 | 5196.1 * |
| Plant Species | Year | Phase | Plant Height | Basal Stem Diameter | Total Leaves | Plant Density |
|---|---|---|---|---|---|---|
| (cm) | (mm) | (No Plant−1) | (Plants m−2) | |||
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |||
| Cyperus papyrus | 2012 | Initial | 33.2 ± 2.4(a) | - | - | 26.0 ± 2.8(b) |
| Final | 46.3 ± 4.5(a) | - | - | 156.0 ± 6.6(a) | ||
| 2013 | Initial | - | - | - | - | |
| Final | - | - | - | - | ||
| Chrysopogon zizanoides | 2012 | Initial | 20.1 ± 1.8(c) | - | - | 5.5 ± 0.0 |
| Final | 251.2 ± 14.8(a) | - | - | 5.5 ± 0.0 | ||
| 2013 | Initial | 58.1 ± 3.7(b) | - | - | 5.5 ± 0.0 | |
| Final | 263.5 ± 3.5(a) | - | - | 5.5 ± 0.0 | ||
| Myscanthus x giganteus | 2012 | Initial | 11.2 ± 0.9(d) | 3.3 ± 0.3(b) | 4.8 ± 1.1(b) | 5.0 ± 1.7(d) |
| Final | 247.3 ± 11.0(b) | 7.1 ± 0.5(a) | 14.8 ± 2.1(a) | 105.0 ± 11.0(b) | ||
| 2013 | Initial | 65.6 ± 3.6(c) | 4.8 ± 0.4(b) | 7.5 ± 2.2(b) | 64.6 ± 3.5(c) | |
| Final | 286.9 ± 21.3(a) | 8.2 ± 0.5(a) | 16.2 ± 1.6(a) | 144.5 ± 6.7(a) | ||
| Arundo donax | 2012 | Initial | 35.1 ± 2.1(c) | 6.1 ± 0.4(c) | 6.2 ± 1.3(c) | 6.0 ± 1.4(c) |
| Final | 437.3 ± 24.5(a) | 23.3 ± 2.5(a) | 42.8 ± 8.9(a) | 30.5 ± 4.9(a) | ||
| 2013 | Initial | 91.9 ± 6.3(b) | 13.3 ± 1.5(b) | 12.1 ± 3.9(b) | 14.4 ± 2.7(b) | |
| Final | 441.5 ± 37.3(a) | 23.9 ± 3.2(a) | 42.9 ± 7.4(a) | 32.0 ± 3.1(a) | ||
| Phragmites australis | 2012 | Initial | 25.4 ± 3.1(c) | 2.7 ± 0.4(b) | 4.3 ± 0.8(b) | 13.3 ± 3.1(c) |
| Final | 248.7 ± 8.1(a) | 8.1 ± 1.1(a) | 17.7 ± 2.1(a) | 364.2 ± 18.6(a) | ||
| 2013 | Initial | 77.3 ± 5.3(b) | 4.6 ± 0.9(b) | 5.9 ± 1.1(b) | 108.1 ± 6.9(b) | |
| Final | 252.6 ± 13.4(a) | 8.7 ± 1.0(a) | 18.3 ± 2.0(a) | 382.0 ± 16.5(a) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milani, M.; Marzo, A.; Toscano, A.; Consoli, S.; Cirelli, G.L.; Ventura, D.; Barbagallo, S. Evapotranspiration from Horizontal Subsurface Flow Constructed Wetlands Planted with Different Perennial Plant Species. Water 2019, 11, 2159. https://doi.org/10.3390/w11102159
Milani M, Marzo A, Toscano A, Consoli S, Cirelli GL, Ventura D, Barbagallo S. Evapotranspiration from Horizontal Subsurface Flow Constructed Wetlands Planted with Different Perennial Plant Species. Water. 2019; 11(10):2159. https://doi.org/10.3390/w11102159
Chicago/Turabian StyleMilani, Mirco, Alessia Marzo, Attilio Toscano, Simona Consoli, Giuseppe Luigi Cirelli, Delia Ventura, and Salvatore Barbagallo. 2019. "Evapotranspiration from Horizontal Subsurface Flow Constructed Wetlands Planted with Different Perennial Plant Species" Water 11, no. 10: 2159. https://doi.org/10.3390/w11102159