
Progranulin Oncogenic Network in Solid Tumors
 , ,
, ,  , , , , ,
, , , , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Progranulin Structure and Nomenclature
3. Progranulin Binding Proteins
3.1. Progranulin Interaction with Extracellular Proteins
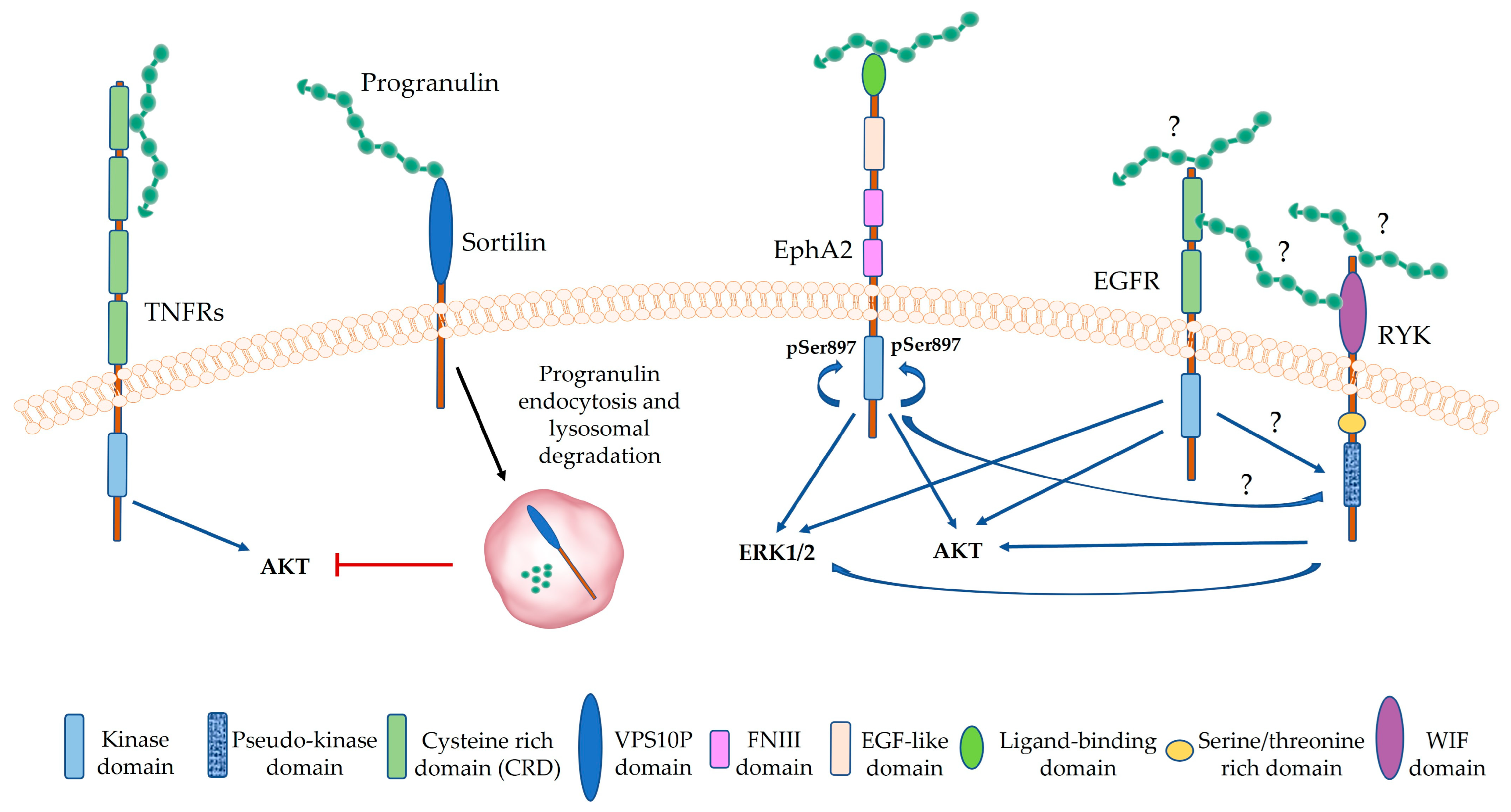
3.2. Progranulin Interaction with Membrane Proteins and Membrane Receptors
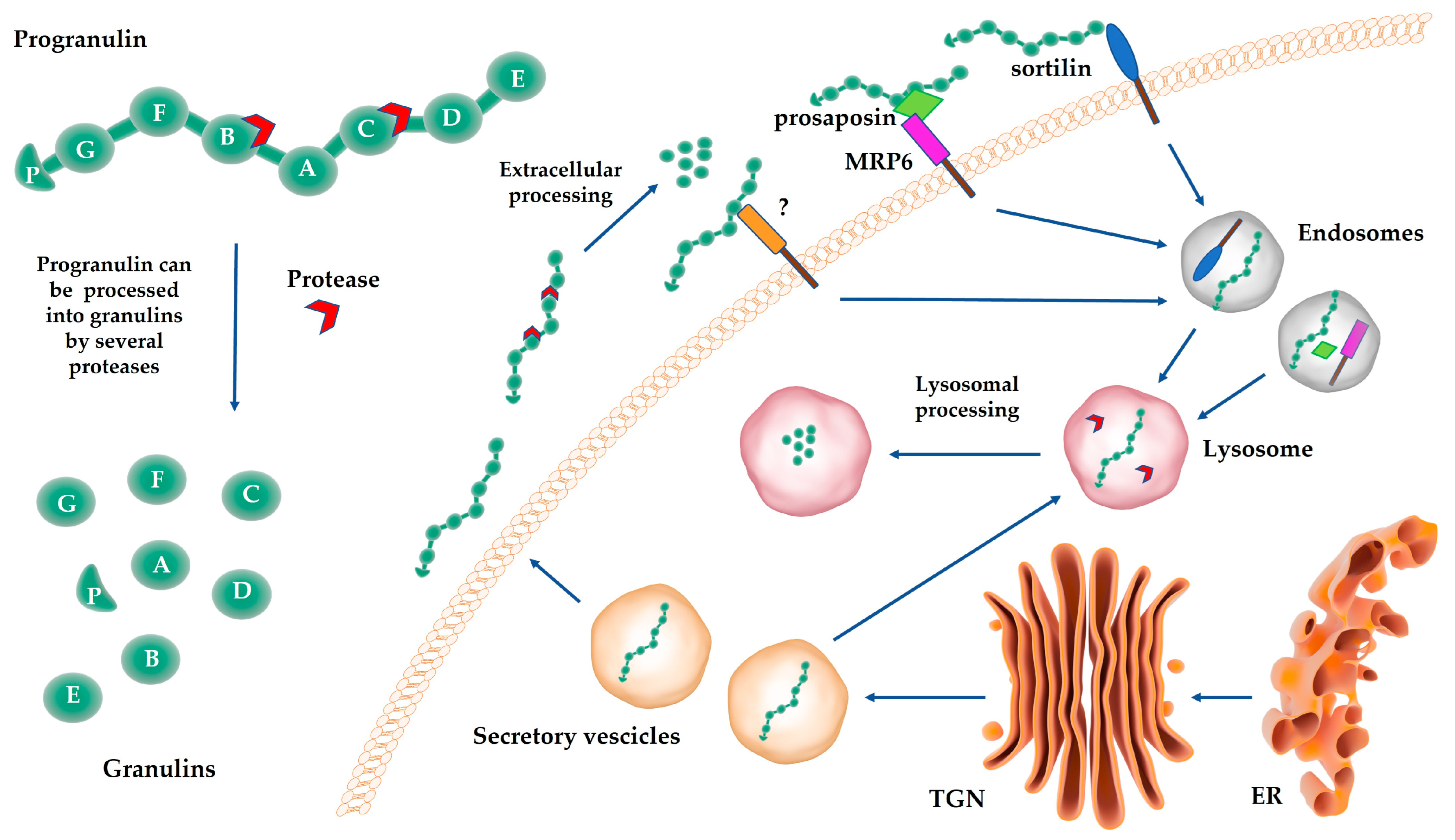
3.3. Progranulin Binding Partners Belonging to the ER/Golgi/Lysosome Network
4. Progranulin in Solid Tumors
4.1. Progranulin Autocrine Function on Tumor Cells
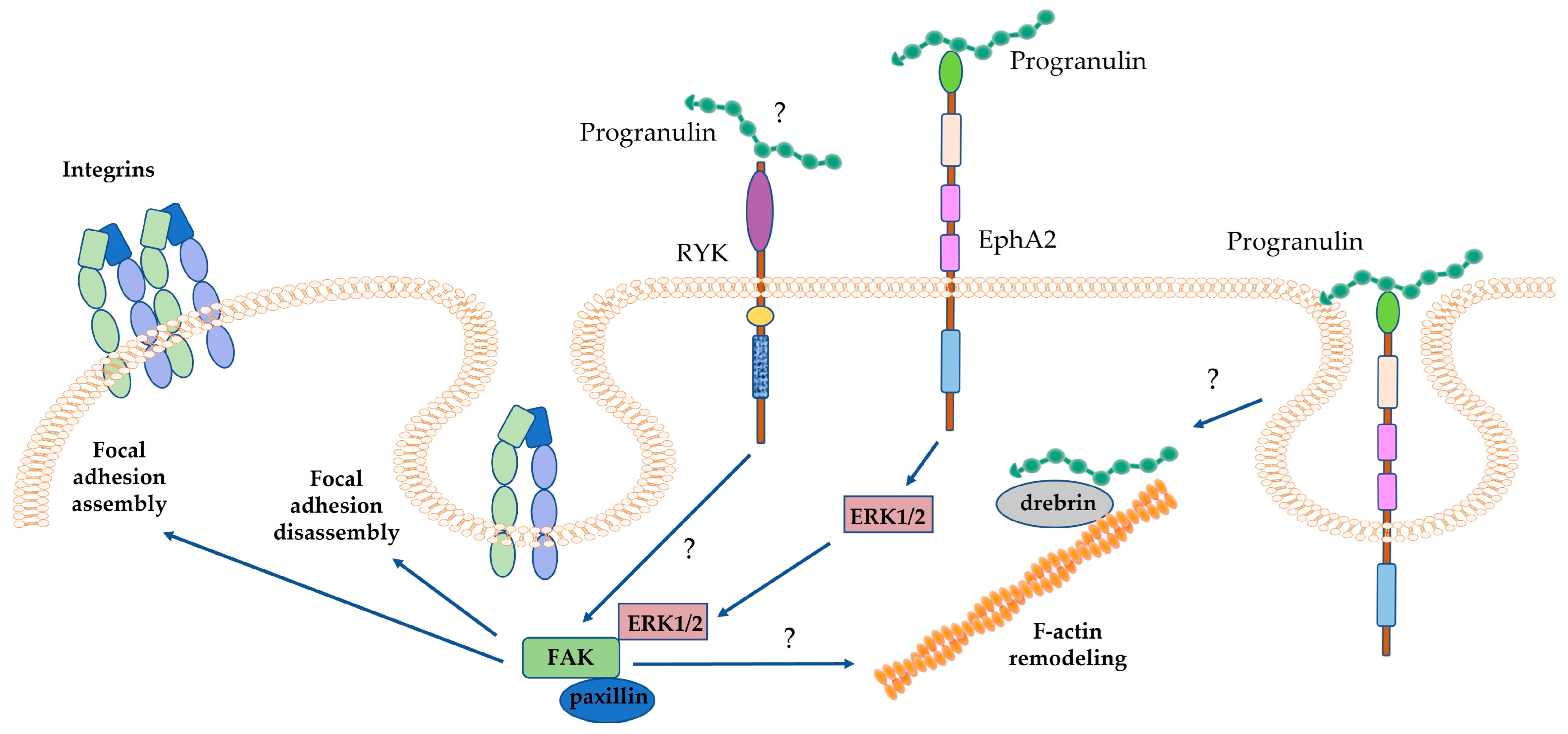
4.1.1. Progranulin and Tumor Cell Proliferation, Migration and Invasion
4.1.2. Progranulin and the Maintenance of CSC
4.2. Progranulin and the Tumor Microenvironment
4.2.1. Progranulin in Tumor Angiogenesis and Lymphangiogenesis
4.2.2. Progranulin and Tumor Immune Evasion
4.2.3. Progranulin and Stromal Fibroblasts/Myofibroblast
4.3. Diagnostic, Prognostic and Predictive Roles of the Progranulin Axis in Cancer
4.4. Progranulin Role in Tumor Resistance to Anticancer Therapies
4.5. Progranulin as a Therapeutic Target in Cancer
5. Progranulin Signaling in Cancer
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diaz-Cueto, L.; Stein, P.; Jacobs, A.; Schultz, R.M.; Gerton, G.L. Modulation of mouse preimplantation embryo development by acrogranin (epithelin/granulin precursor). Dev. Biol. 2000, 217, 406–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Diaz-Cueto, L.; Schwarze, J.E.; Takahashi, Y.; Imai, M.; Isuzugawa, K.; Yamamoto, S.; Chang, K.T.; Gerton, G.L.; Imakawa, K. Effects of progranulin on blastocyst hatching and subsequent adhesion and outgrowth in the mouse. Biol. Reprod. 2005, 73, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Desmarais, J.A.; Cao, M.; Bateman, A.; Murphy, B.D. Spatiotemporal expression pattern of progranulin in embryo implantation and placenta formation suggests a role in cell proliferation, remodeling, and angiogenesis. Reproduction 2008, 136, 247–257. [Google Scholar] [CrossRef]
- He, Z.; Bateman, A. Progranulin (granulin-epithelin precursor, PC-cell-derived growth factor, acrogranin) mediates tissue repair and tumorigenesis. J. Mol. Med. 2003, 81, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.P.; Tian, Q.Y.; Frenkel, S.; Liu, C.J. The promotion of bone healing by progranulin, a downstream molecule of BMP-2, through interacting with TNF/TNFR signaling. Biomaterials 2013, 34, 6412–6421. [Google Scholar] [CrossRef] [Green Version]
- Toh, H.; Cao, M.; Daniels, E.; Bateman, A. Expression of the growth factor progranulin in endothelial cells influences growth and development of blood vessels: A novel mouse model. PLoS ONE 2013, 8, e64989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Lu, Y.; Tian, Q.Y.; Zhang, Y.; Guo, F.J.; Liu, G.Y.; Syed, N.M.; Lai, Y.; Lin, E.A.; Kong, L.; et al. The growth factor progranulin binds to TNF receptors and is therapeutic against inflammatory arthritis in mice. Science 2011, 332, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Yin, F.; Banerjee, R.; Thomas, B.; Zhou, P.; Qian, L.; Jia, T.; Ma, X.; Ma, Y.; Iadecola, C.; Beal, M.F.; et al. Exaggerated inflammation, impaired host defense, and neuropathology in progranulin-deficient mice. J. Exp. Med. 2010, 207, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Park, B.; Buti, L.; Lee, S.; Matsuwaki, T.; Spooner, E.; Brinkmann, M.M.; Nishihara, M.; Ploegh, H.L. Granulin is a soluble cofactor for toll-like receptor 9 signaling. Immunity 2011, 34, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, P.; Van Hoecke, A.; Lambrechts, D.; Vanacker, P.; Bogaert, E.; van Swieten, J.; Carmeliet, P.; Van Den Bosch, L.; Robberecht, W. Progranulin functions as a neurotrophic factor to regulate neurite outgrowth and enhance neuronal survival. J. Cell Biol. 2008, 181, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Ryan, C.L.; Baranowski, D.C.; Chitramuthu, B.P.; Malik, S.; Li, Z.; Cao, M.; Minotti, S.; Durham, H.D.; Kay, D.G.; Shaw, C.A.; et al. Progranulin is expressed within motor neurons and promotes neuronal cell survival. BMC Neurosci. 2009, 10, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, L.H.; Zhang, J.; Barmada, S.J.; Zhou, P.; Kamiya, S.; Sun, B.; Min, S.W.; Gan, L.; Finkbeiner, S.; Huang, E.J.; et al. Progranulin deficiency promotes neuroinflammation and neuron loss following toxin-induced injury. J. Clin. Investig. 2022, 132, 157161. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Padukkavidana, T.; Vaegter, C.B.; Brady, O.A.; Zheng, Y.; Mackenzie, I.R.; Feldman, H.H.; Nykjaer, A.; Strittmatter, S.M. Sortilin-mediated endocytosis determines levels of the frontotemporal dementia protein, progranulin. Neuron 2010, 68, 654–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Matsuwaki, T.; Yamanouchi, K.; Nishihara, M. Increased lysosomal biogenesis in activated microglia and exacerbated neuronal damage after traumatic brain injury in progranulin-deficient mice. Neuroscience 2013, 250, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Cheung, S.T.; Bennett, H.P.J. A Brief Overview of Progranulin in Health and Disease. Methods Mol. Biol. 2018, 1806, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Hettinghouse, A.; Liu, C.J. Progranulin: A conductor of receptors orchestra, a chaperone of lysosomal enzymes and a therapeutic target for multiple diseases. Cytokine Growth Factor Rev. 2019, 45, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Chitramuthu, B.P.; Bennett, H.P.J.; Bateman, A. Progranulin: A new avenue towards the understanding and treatment of neurodegenerative disease. Brain 2017, 140, 3081–3104. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.; Mackenzie, I.R.; Pickering-Brown, S.M.; Gass, J.; Rademakers, R.; Lindholm, C.; Snowden, J.; Adamson, J.; Sadovnick, A.D.; Rollinson, S.; et al. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature 2006, 24, 916–919. [Google Scholar] [CrossRef]
- Cruts, M.; Gijselinck, I.; van der Zee, J.; Engelborghs, S.; Wils, H.; Pirici, D.; Rademakers, R.; Vandenberghe, R.; Dermaut, B.; Martin, J.J.; et al. Null mutations in progranulin cause ubiquitin-positive frontotemporal dementia linked to chromosome 17q21. Nature 2006, 442, 920–924. [Google Scholar] [CrossRef]
- Jian, J.; Li, G.; Hettinghouse, A.; Liu, C. Progranulin: A key player in autoimmune diseases. Cytokine 2018, 101, 48–55. [Google Scholar] [CrossRef]
- Palfree, R.G.; Bennett, H.P.; Bateman, A. The Evolution of the Secreted Regulatory Protein Progranulin. PLoS ONE 2015, 10, e0133749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolkatchev, D.; Malik, S.; Vinogradova, A.; Wang, P.; Chen, Z.; Xu, P.; Bennett, H.P.; Bateman, A.; Ni, F. Structure dissection of human progranulin identifies well-folded granulin/epithelin modules with unique functional activities. Protein Sci. 2008, 17, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrabal, R.; Chen, Z.; James, S.; Bennett, H.P.; Ni, F. The hairpin stack fold, a novel protein architecture for a new family of protein growth factors. Nat. Struct. Biol. 1996, 3, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Petoukhov, E.; Fernando, S.; Mills, F.; Shivji, F.; Hunter, D.; Krieger, C.; Silverman, M.A.; Bamji, S.X. Activity-dependent secretion of progranulin from synapses. J. Cell Sci. 2013, 126, 5412–5421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benussi, L.; Ciani, M.; Tonoli, E.; Morbin, M.; Palamara, L.; Albani, D.; Fusco, F.; Forloni, G.; Glionna, M.; Baco, M.; et al. Loss of exosomes in progranulin-associated frontotemporal dementia. Neurobiol. Aging 2016, 40, 41–49. [Google Scholar] [CrossRef]
- Songsrirote, K.; Li, Z.; Ashford, D.; Bateman, A.; Thomas-Oates, J. Development and application of mass spectrometric methods for the analysis of progranulin N-glycosylation. J. Proteom. 2010, 73, 1479–1490. [Google Scholar] [CrossRef]
- Suh, H.S.; Choi, N.; Tarassishin, L.; Lee, S.C. Regulation of progranulin expression in human microglia and proteolysis of progranulin by matrix metalloproteinase-12 (MMP-12). PLoS ONE 2012, 7, e35115. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Nathan, C.; Jin, W.; Sim, D.; Ashcroft, G.S.; Wahl, S.M.; Lacomis, L.; Erdjument-Bromage, H.; Tempst, P.; Wright, C.D.; et al. Conversion of proepithelin to epithelins: Roles of SLPI and elastase in host defense and wound repair. Cell 2002, 111, 867–878. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Frohlich, L.; Sixt, M.; Lammermann, T.; Pfister, H.; Bateman, A.; Belaaouaj, A.; Ring, J.; Ollert, M.; Fassler, R.; et al. Proteinase 3 and neutrophil elastase enhance inflammation in mice by inactivating antiinflammatory progranulin. J. Clin. Investig. 2008, 118, 2438–2447. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Lai, Y.; Tian, Q.; Lin, E.A.; Kong, L.; Liu, C. Granulin-epithelin precursor binds directly to ADAMTS-7 and ADAMTS-12 and inhibits their degradation of cartilage oligomeric matrix protein. Arthritis Rheum. 2010, 62, 2023–2036. [Google Scholar] [CrossRef]
- Okura, H.; Yamashita, S.; Ohama, T.; Saga, A.; Yamamoto-Kakuta, A.; Hamada, Y.; Sougawa, N.; Ohyama, R.; Sawa, Y.; Matsuyama, A. HDL/apolipoprotein A-I binds to macrophage-derived progranulin and suppresses its conversion into proinflammatory granulins. J. Atheroscler. Thromb. 2010, 17, 568–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.; Bennett, H.P. The granulin gene family: From cancer to dementia. Bioessays 2009, 31, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Parnell, P.G.; Wunderlich, J.; Carter, B.; Halper, J. Transforming growth factor e: Amino acid analysis and partial amino acid sequence. Growth Factors 1992, 7, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Sun, L.; Bastos de Oliveira, F.; Qi, X.; Brown, W.J.; Smolka, M.B.; Sun, Y.; Hu, F. Prosaposin facilitates sortilin-independent lysosomal trafficking of progranulin. J. Cell Biol. 2015, 210, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Holler, C.J.; Taylor, G.; Deng, Q.; Kukar, T. Intracellular Proteolysis of Progranulin Generates Stable, Lysosomal Granulins that Are Haploinsufficient in Patients with Frontotemporal Dementia Caused by GRN Mutations. eNeuro 2017, 4, 5562298. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Stankowski, J.N.; Chew, J.; Cook, C.N.; Lam, Y.W.; Almeida, S.; Carlomagno, Y.; Lau, K.F.; Prudencio, M.; Gao, F.B.; et al. The lysosomal protein cathepsin L is a progranulin protease. Mol. Neurodegener. 2017, 12, 55. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A.; Belcourt, D.; Bennett, H.; Lazure, C.; Solomon, S. Granulins, a novel class of peptide from leukocytes. Biochem. Biophys. Res. Commun. 1990, 173, 1161–1168. [Google Scholar] [CrossRef]
- Shoyab, M.; McDonald, V.L.; Byles, C.; Todaro, G.J.; Plowman, G.D. Epithelins 1 and 2: Isolation and characterization of two cysteine-rich growth-modulating proteins. Proc. Natl. Acad. Sci. USA 1990, 87, 7912–7916. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, V.; Palfree, R.G.; Bateman, A. Isolation and sequence of the granulin precursor cDNA from human bone marrow reveals tandem cysteine-rich granulin domains. Proc. Natl. Acad. Sci. USA 1992, 89, 1715–1719. [Google Scholar] [CrossRef] [Green Version]
- Plowman, G.D.; Green, J.M.; Neubauer, M.G.; Buckley, S.D.; McDonald, V.L.; Todaro, G.J.; Shoyab, M. The epithelin precursor encodes two proteins with opposing activities on epithelial cell growth. J. Biol. Chem. 1992, 267, 13073–13078. [Google Scholar] [CrossRef]
- Anakwe, O.O.; Gerton, G.L. Acrosome biogenesis begins during meiosis: Evidence from the synthesis and distribution of an acrosomal glycoprotein, acrogranin, during guinea pig spermatogenesis. Biol. Reprod. 1990, 42, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Gao, G.; Crabb, J.W.; Serrero, G. Purification of an autocrine growth factor homologous with mouse epithelin precursor from a highly tumorigenic cell line. J. Biol. Chem. 1993, 268, 10863–10869. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Serrero, G. Inhibition of PC cell-derived growth factor (PCDGF, epithelin/granulin precursor) expression by antisense PCDGF cDNA transfection inhibits tumorigenicity of the human breast carcinoma cell line MDA-MB-468. Proc. Natl. Acad. Sci. USA 2000, 97, 3993–3998. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Serrero, G. Mediation of estrogen mitogenic effect in human breast cancer MCF-7 cells by PC-cell-derived growth factor (PCDGF/granulin precursor). Proc. Natl. Acad. Sci. USA 2001, 98, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Serrero, G. Identification of cell surface binding sites for PC-cell-derived growth factor, PCDGF, (epithelin/granulin precursor) on epithelial cells and fibroblasts. Biochem. Biophys. Res. Commun. 1998, 245, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Serrero, G. Inhibition of tumorigenicity of the teratoma PC cell line by transfection with antisense cDNA for PC cell-derived growth factor (PCDGF, epithelin/granulin precursor). Proc. Natl. Acad. Sci. USA 1998, 95, 14202–14207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, T.; Hoff, H.B., III; Nemoto, H.; Lee, H.; Orth, J.; Arai, Y.; Gerton, G.L. Acrogranin, an acrosomal cysteine-rich glycoprotein, is the precursor of the growth-modulating peptides, granulins, and epithelins, and is expressed in somatic as well as male germ cells. Mol. Reprod. Dev. 1993, 34, 233–243. [Google Scholar] [CrossRef]
- Chitramuthu, B.P.; Campos-García, V.R.; Bateman, A. Multiple Molecular Pathways Are Influenced by Progranulin in a Neuronal Cell Model-A Parallel Omics Approach. Front. Neurosci. 2021, 15, 775391. [Google Scholar] [CrossRef]
- Hoque, M.; Tian, B.; Mathews, M.B.; Pe’ery, T. Granulin and granulin repeats interact with the Tat.P-TEFb complex and inhibit Tat transactivation. J. Biol. Chem. 2005, 280, 13648–13657. [Google Scholar] [CrossRef] [Green Version]
- Hoque, M.; Young, T.M.; Lee, C.G.; Serrero, G.; Mathews, M.B.; Pe’ery, T. The growth factor granulin interacts with cyclin T1 and modulates P-TEFb-dependent transcription. Mol. Cell Biol. 2003, 23, 1688–1702. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, E.M.; Mongiat, M.; Slater, S.J.; Baffa, R.; Iozzo, R.V. A novel interaction between perlecan protein core and progranulin: Potential effects on tumor growth. J. Biol Chem. 2003, 278, 38113–38116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abella, V.; Pino, J.; Scotece, M.; Conde, J.; Lago, F.; Gonzalez-Gay, M.A.; Mera, A.; Gomez, R.; Mobasheri, A.; Gualillo, O. Progranulin as a biomarker and potential therapeutic agent. Drug Discov. Today 2017, 22, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Zhang, Y.; Ilalov, K.; Carlson, C.S.; Feng, J.Q.; Di Cesare, P.E.; Liu, C.J. Cartilage oligomeric matrix protein associates with granulin-epithelin precursor (GEP) and potentiates GEP-stimulated chondrocyte proliferation. J. Biol. Chem. 2007, 282, 11347–11355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Zhao, Y.P.; Tian, Q.Y.; Feng, J.Q.; Kobayashi, T.; Merregaert, J.; Liu, C.J. Extracellular matrix protein 1, a direct targeting molecule of parathyroid hormone-related peptide, negatively regulates chondrogenesis and endochondral ossification via associating with progranulin growth factor. FASEB J. 2016, 30, 2741–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, A.M.; Finch, N.A.; Almeida, M.; Perkerson, R.B.; van Blitterswijk, M.; Wojtas, A.; Cenik, B.; Rotondo, S.; Inskeep, V.; Almasy, L.; et al. Prosaposin is a regulator of progranulin levels and oligomerization. Nat. Commun. 2016, 7, 11992. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Li, X.X.; Gao, W.; Liu, W.; Liu, D.S. Progranulin-derived Atsttrin directly binds to TNFRSF25 (DR3) and inhibits TNF-like ligand 1A (TL1A) activity. PLoS ONE 2014, 9, e92743. [Google Scholar] [CrossRef] [Green Version]
- Altmann, C.; Vasic, V.; Hardt, S.; Heidler, J.; Häussler, A.; Wittig, I.; Schmidt, M.H.H.; Tegeder, I. Progranulin promotes peripheral nerve regeneration and reinnervation: Role of notch signaling. Mol. Neurodegener. 2016, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Baladron, V.; Ruiz-Hidalgo, M.J.; Bonvini, E.; Gubina, E.; Notario, V.; Laborda, J. The EGF-like homeotic protein dlk affects cell growth and interacts with growth-modulating molecules in the yeast two-hybrid system. Biochem. Biophys. Res. Commun. 2002, 291, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Neill, T.; Buraschi, S.; Goyal, A.; Sharpe, C.; Natkanski, E.; Schaefer, L.; Morrione, A.; Iozzo, R.V. EphA2 is a functional receptor for the growth factor progranulin. J. Cell Biol. 2016, 215, 687–703. [Google Scholar] [CrossRef] [Green Version]
- Terryn, J.; Verfaillie, C.M.; Van Damme, P. Tweaking Progranulin Expression: Therapeutic Avenues and Opportunities. Front. Mol. Neurosci. 2021, 14, 713031. [Google Scholar] [CrossRef]
- Etemadi, N.; Webb, A.; Bankovacki, A.; Silke, J.; Nachbur, U. Progranulin does not inhibit TNF and lymphotoxin-alpha signalling through TNF receptor 1. Immunol. Cell Biol. 2013, 91, 661–664. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chang, J.; Deng, Q.; Xu, J.; Nguyen, T.A.; Martens, L.H.; Cenik, B.; Taylor, G.; Hudson, K.F.; Chung, J.; et al. Progranulin does not bind tumor necrosis factor (TNF) receptors and is not a direct regulator of TNF-dependent signaling or bioactivity in immune or neuronal cells. J. Neurosci. 2013, 33, 9202–9213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, I.; Fullsack, S.; Wajant, H. Lack of Evidence for a Direct Interaction of Progranulin and Tumor Necrosis Factor Receptor-1 and Tumor Necrosis Factor Receptor-2 From Cellular Binding Studies. Front. Immunol. 2018, 9, 793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.C.; Liu, H.; Talwar, A.; Jian, J. New discovery rarely runs smooth: An update on progranulin/TNFR interactions. Protein Cell 2015, 6, 792–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, E.; Xie, C.; Buraschi, S.; Belfiore, A.; Iozzo, R.V.; Giordano, A.; Morrione, A. Complexity of progranulin mechanisms of action in mesothelioma. J. Exp. Clin. Cancer Res. 2022, 41, 333. [Google Scholar] [CrossRef]
- Zheng, Y.; Brady, O.A.; Meng, P.S.; Mao, Y.; Hu, F. C-terminus of progranulin interacts with the beta-propeller region of sortilin to regulate progranulin trafficking. PLoS ONE 2011, 6, e21023. [Google Scholar] [CrossRef]
- Zhou, X.; Sullivan, P.M.; Sun, L.; Hu, F. The interaction between progranulin and prosaposin is mediated by granulins and the linker region between saposin B and C. J. Neurochem. 2017, 143, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Jian, J.; Zhao, S.; Tian, Q.; Gonzalez-Gugel, E.; Mundra, J.J.; Uddin, S.M.; Liu, B.; Richbourgh, B.; Brunetti, R.; Liu, C.J. Progranulin directly binds to the CRD2 and CRD3 of TNFR extracellular domains. FEBS Lett. 2013, 587, 3428–3436. [Google Scholar] [CrossRef] [Green Version]
- Paushter, D.H.; Du, H.; Feng, T.; Hu, F. The lysosomal function of progranulin, a guardian against neurodegeneration. Acta Neuropathol. 2018, 136, 1–17. [Google Scholar] [CrossRef]
- He, Z.; Bateman, A. Progranulin gene expression regulates epithelial cell growth and promotes tumor growth in vivo. Cancer Res. 1999, 59, 3222–3229. [Google Scholar]
- He, Z.; Ismail, A.; Kriazhev, L.; Sadvakassova, G.; Bateman, A. Progranulin (PC-cell-derived growth factor/acrogranin) regulates invasion and cell survival. Cancer Res. 2002, 62, 5590–5596. [Google Scholar]
- He, Z.; Ong, C.H.; Halper, J.; Bateman, A. Progranulin is a mediator of the wound response. Nat. Med. 2003, 9, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Serrero, G. Progranulin/GP88, A Complex and Multifaceted Player of Tumor Growth by Direct Action and via the Tumor Microenvironment. Adv. Exp. Med. Biol. 2021, 1329, 475–498. [Google Scholar] [CrossRef] [PubMed]
- Arechavaleta-Velasco, F.; Perez-Juarez, C.E.; Gerton, G.L.; Diaz-Cueto, L. Progranulin and its biological effects in cancer. Med. Oncol. 2017, 34, 194. [Google Scholar] [CrossRef]
- Daya, M.; Loilome, W.; Techasen, A.; Thanee, M.; Sa-Ngiamwibool, P.; Titapun, A.; Yongvanit, P.; Namwat, N. Progranulin modulates cholangiocarcinoma cell proliferation, apoptosis, and motility via the PI3K/pAkt pathway. Onco Targets Ther. 2018, 11, 395–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, R.; Yue, B.; Dong, J.; Banerjee, A.; Serrero, G. Anti-progranulin/GP88 antibody AG01 inhibits triple negative breast cancer cell proliferation and migration. Breast Cancer Res. Treat. 2021, 186, 637–653. [Google Scholar] [CrossRef]
- Yabe, K.; Yamamoto, Y.; Takemura, M.; Hara, T.; Tsurumi, H.; Serrero, G.; Nabeshima, T.; Saito, K. Progranulin depletion inhibits proliferation via the transforming growth factor beta/SMAD family member 2 signaling axis in Kasumi-1 cells. Heliyon 2021, 7, e05849. [Google Scholar] [CrossRef]
- Monami, G.; Gonzalez, E.M.; Hellman, M.; Gomella, L.G.; Baffa, R.; Iozzo, R.V.; Morrione, A. Proepithelin promotes migration and invasion of 5637 bladder cancer cells through the activation of ERK1/2 and the formation of a paxillin/FAK/ERK complex. Cancer Res. 2006, 66, 7103–7110. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.Q.; Buraschi, S.; Morcavallo, A.; Genua, M.; Shirao, T.; Peiper, S.C.; Gomella, L.G.; Birbe, R.; Belfiore, A.; Iozzo, R.V.; et al. A novel role for drebrin in regulating progranulin bioactivity in bladder cancer. Oncotarget 2015, 6, 10825–10839. [Google Scholar] [CrossRef] [Green Version]
- Walcher, L.; Kistenmacher, A.K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauss, A.; Blaudszun, A.R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Cheung, P.F.; Cheng, C.K.; Wong, N.C.; Ho, J.C.; Yip, C.W.; Lui, V.C.; Cheung, A.N.; Fan, S.T.; Cheung, S.T. Granulin-epithelin precursor is an oncofetal protein defining hepatic cancer stem cells. PLoS ONE 2011, 6, e28246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandey, I.; Chiou, S.H.; Huang, A.P.; Tsai, J.C.; Tu, P.H. Progranulin promotes Temozolomide resistance of glioblastoma by orchestrating DNA repair and tumor stemness. Oncogene 2015, 34, 1853–1864. [Google Scholar] [CrossRef] [PubMed]
- Rhost, S.; Hughes, E.; Harrison, H.; Rafnsdottir, S.; Jacobsson, H.; Gregersson, P.; Magnusson, Y.; Fitzpatrick, P.; Andersson, D.; Berger, K.; et al. Sortilin inhibition limits secretion-induced progranulin-dependent breast cancer progression and cancer stem cell expansion. Breast Cancer Res. 2018, 20, 137. [Google Scholar] [CrossRef] [Green Version]
- Berger, K.; Pauwels, E.; Parkinson, G.; Landberg, G.; Le, T.; Demillo, V.G.; Lumangtad, L.A.; Jones, D.E.; Islam, M.A.; Olsen, R.; et al. Reduction of Progranulin-Induced Breast Cancer Stem Cell Propagation by Sortilin-Targeting Cyclotriazadisulfonamide (CADA) Compounds. J. Med. Chem. 2021, 64, 12865–12876. [Google Scholar] [CrossRef]
- Davidson, B.; Alejandro, E.; Florenes, V.A.; Goderstad, J.M.; Risberg, B.; Kristensen, G.B.; Trope, C.G.; Kohn, E.C. Granulin-epithelin precursor is a novel prognostic marker in epithelial ovarian carcinoma. Cancer 2004, 100, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Li, J.S.; Liang, Q.P.; He, D.Z.; Zhao, J. Expression of PC cell-derived growth factor and vascular endothelial growth factor in esophageal squamous cell carcinoma and their clinicopathologic significance. Chin. Med. J. (Engl.) 2008, 121, 881–886. [Google Scholar] [CrossRef]
- Wang, M.; Li, G.; Yin, J.; Lin, T.; Zhang, J. Progranulin overexpression predicts overall survival in patients with glioblastoma. Med. Oncol. 2012, 29, 2423–2431. [Google Scholar] [CrossRef]
- Yang, D.; Wang, L.L.; Dong, T.T.; Shen, Y.H.; Guo, X.S.; Liu, C.Y.; Liu, J.; Zhang, P.; Li, J.; Sun, Y.P. Progranulin promotes colorectal cancer proliferation and angiogenesis through TNFR2/Akt and ERK signaling pathways. Am. J. Cancer Res. 2015, 5, 3085–3097. [Google Scholar]
- Tangkeangsirisin, W.; Serrero, G. PC cell-derived growth factor (PCDGF/GP88, progranulin) stimulates migration, invasiveness and VEGF expression in breast cancer cells. Carcinogenesis 2004, 25, 1587–1592. [Google Scholar] [CrossRef]
- Li, L.Q.; Huang, H.L.; Ping, J.L.; Wang, X.H.; Zhong, J.; Dai, L.C. Clinicopathologic and prognostic implications of progranulin in breast carcinoma. Chin. Med. J. (Engl.) 2011, 124, 2045–2050. [Google Scholar]
- Eguchi, R.; Nakano, T.; Wakabayashi, I. Progranulin and granulin-like protein as novel VEGF-independent angiogenic factors derived from human mesothelioma cells. Oncogene 2017, 36, 714–722. [Google Scholar] [CrossRef]
- Huang, H.; Li, J.; Lu, Y.; Min, L.; Li, D.; Dai, L. Role of midkine-progranulin interaction during angiogenesis of hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 8809–8820. [Google Scholar]
- Li, G.; Dong, T.; Yang, D.; Gao, A.; Luo, J.; Yang, H.; Wang, L. Progranulin promotes lymphangiogenesis through VEGF-C and is an independent risk factor in human esophageal cancers. Hum. Pathol. 2018, 75, 116–124. [Google Scholar] [CrossRef]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198. [Google Scholar] [CrossRef]
- Kwack, K.H.; Lee, H.W. Progranulin Inhibits Human T Lymphocyte Proliferation by Inducing the Formation of Regulatory T Lymphocytes. Mediat. Inflamm. 2017, 2017, 7682083. [Google Scholar] [CrossRef] [Green Version]
- Cheung, P.F.; Yip, C.W.; Wong, N.C.; Fong, D.Y.; Ng, L.W.; Wan, A.M.; Wong, C.K.; Cheung, T.T.; Ng, I.O.; Poon, R.T.; et al. Granulin-epithelin precursor renders hepatocellular carcinoma cells resistant to natural killer cytotoxicity. Cancer Immunol. Res. 2014, 2, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Cheung, P.F.; Yip, C.W.; Ng, L.W.; Wong, C.K.; Cheung, T.T.; Lo, C.M.; Fan, S.T.; Cheung, S.T. Restoration of natural killer activity in hepatocellular carcinoma by treatment with antibody against granulin-epithelin precursor. Oncoimmunology 2015, 4, e1016706. [Google Scholar] [CrossRef] [Green Version]
- Quaranta, V.; Rainer, C.; Nielsen, S.R.; Raymant, M.L.; Ahmed, M.S.; Engle, D.D.; Taylor, A.; Murray, T.; Campbell, F.; Palmer, D.H.; et al. Macrophage-Derived Granulin Drives Resistance to Immune Checkpoint Inhibition in Metastatic Pancreatic Cancer. Cancer Res. 2018, 78, 4253–4269. [Google Scholar] [CrossRef] [Green Version]
- Voshtani, R.; Song, M.; Wang, H.; Li, X.; Zhang, W.; Tavallaie, M.S.; Yan, W.; Sun, J.; Wei, F.; Ma, X. Progranulin promotes melanoma progression by inhibiting natural killer cell recruitment to the tumor microenvironment. Cancer Lett. 2019, 465, 24–35. [Google Scholar] [CrossRef]
- Fang, W.; Zhou, T.; Shi, H.; Yao, M.; Zhang, D.; Qian, H.; Zeng, Q.; Wang, Y.; Jin, F.; Chai, C.; et al. Progranulin induces immune escape in breast cancer via up-regulating PD-L1 expression on tumor-associated macrophages (TAMs) and promoting CD8(+) T cell exclusion. J. Exp. Clin. Cancer Res. 2021, 40, 4. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Ye, X.; Zhou, T.; Gan, D.; Qian, H.; Fang, W.; Yao, M.; Zhang, D.; Shi, H.; Chen, T. PGRN(-/-) TAMs-derived exosomes inhibit breast cancer cell invasion and migration and its mechanism exploration. Life Sci. 2021, 264, 118687. [Google Scholar] [CrossRef] [PubMed]
- Cheung, P.F.; Yang, J.; Fang, R.; Borgers, A.; Krengel, K.; Stoffel, A.; Althoff, K.; Yip, C.W.; Siu, E.H.L.; Ng, L.W.C.; et al. Progranulin mediates immune evasion of pancreatic ductal adenocarcinoma through regulation of MHCI expression. Nat. Commun. 2022, 13, 156. [Google Scholar] [CrossRef]
- Elkabets, M.; Gifford, A.M.; Scheel, C.; Nilsson, B.; Reinhardt, F.; Bray, M.A.; Carpenter, A.E.; Jirstrom, K.; Magnusson, K.; Ebert, B.L.; et al. Human tumors instigate granulin-expressing hematopoietic cells that promote malignancy by activating stromal fibroblasts in mice. J. Clin. Investig. 2011, 121, 784–799. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.R.; Quaranta, V.; Linford, A.; Emeagi, P.; Rainer, C.; Santos, A.; Ireland, L.; Sakai, T.; Sakai, K.; Kim, Y.S.; et al. Macrophage-secreted granulin supports pancreatic cancer metastasis by inducing liver fibrosis. Nat. Cell Biol. 2016, 18, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, D.; Tian, J.; Gao, A.; Shen, Y.; Ren, X.; Li, X.; Jiang, G.; Dong, T. Tumor necrosis factor receptor 2/AKT and ERK signaling pathways contribute to the switch from fibroblasts to CAFs by progranulin in microenvironment of colorectal cancer. Oncotarget 2017, 8, 26323–26333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.E.; Serrero, G. PC cell-derived growth factor stimulates proliferation and confers Trastuzumab resistance to Her-2-overexpressing breast cancer cells. Clin. Cancer Res. 2006, 12, 4192–4199. [Google Scholar] [CrossRef] [Green Version]
- Serrero, G. Autocrine growth factor revisited: PC-cell-derived growth factor (progranulin), a critical player in breast cancer tumorigenesis. Biochem. Biophys. Res. Commun. 2003, 308, 409–413. [Google Scholar] [CrossRef]
- Serrero, G.; Hawkins, D.M.; Yue, B.; Ioffe, O.; Bejarano, P.; Phillips, J.T.; Head, J.F.; Elliott, R.L.; Tkaczuk, K.R.; Godwin, A.K.; et al. Progranulin (GP88) tumor tissue expression is associated with increased risk of recurrence in breast cancer patients diagnosed with estrogen receptor positive invasive ductal carcinoma. Breast Cancer Res. 2012, 14, R26. [Google Scholar] [CrossRef] [Green Version]
- Serrero, G.; Hicks, D. Immunohistochemical Detection of Progranulin (PGRN/GP88/GEP) in Tumor Tissues as a Cancer Prognostic Biomarker. Methods Mol. Biol. 2018, 1806, 107–120. [Google Scholar] [CrossRef]
- Serrero, G.; Ioffe, O.B. Expression of PC-cell-derived growth factor in benign and malignant human breast epithelium. Hum. Pathol. 2003, 34, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Tkaczuk, K.H.R.; Hawkins, D.; Yue, B.; Hicks, D.; Tait, N.; Serrero, G. Association of Serum Progranulin Levels With Disease Progression, Therapy Response and Survival in Patients With Metastatic Breast Cancer. Clin. Breast Cancer 2019, 20, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkaczuk, K.R.; Yue, B.; Zhan, M.; Tait, N.; Yarlagadda, L.; Dai, H.; Serrero, G. Increased Circulating Level of the Survival Factor GP88 (Progranulin) in the Serum of Breast Cancer Patients When Compared to Healthy Subjects. Breast Cancer (Auckl.) 2011, 5, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Koo, D.H.; Park, C.Y.; Lee, E.S.; Ro, J.; Oh, S.W. Progranulin as a prognostic biomarker for breast cancer recurrence in patients who had hormone receptor-positive tumors: A cohort study. PLoS ONE 2012, 7, e39880. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.; Rhost, S.; Rafnsdottir, S.; Hughes, E.; Magnusson, Y.; Ekholm, M.; Stal, O.; Ryden, L.; Landberg, G. Tumor co-expression of progranulin and sortilin as a prognostic biomarker in breast cancer. BMC Cancer 2021, 21, 185. [Google Scholar] [CrossRef]
- Pan, C.X.; Kinch, M.S.; Kiener, P.A.; Langermann, S.; Serrero, G.; Sun, L.; Corvera, J.; Sweeney, C.J.; Li, L.; Zhang, S.; et al. PC cell-derived growth factor expression in prostatic intraepithelial neoplasia and prostatic adenocarcinoma. Clin. Cancer Res. 2004, 10, 1333–1337. [Google Scholar] [CrossRef] [Green Version]
- Monami, G.; Emiliozzi, V.; Bitto, A.; Lovat, F.; Xu, S.Q.; Goldoni, S.; Fassan, M.; Serrero, G.; Gomella, L.G.; Baffa, R.; et al. Proepithelin regulates prostate cancer cell biology by promoting cell growth, migration, and anchorage-independent growth. Am. J. Pathol. 2009, 174, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Greither, T.; Fischer, K.; Theil, G.; Marcou, M.; Holzhausen, H.J.; Weigelt, K.; Serrero, G.; Hicks, D.; Yue, B.; Fornara, P.; et al. Expression of GP88 (progranulin) in serum of prostate cancer patients is associated with Gleason scores and overall survival. Cancer Manag. Res. 2018, 10, 4173–4180. [Google Scholar] [CrossRef] [Green Version]
- Fichte, A.; Neumann, A.; Weigelt, K.; Guzman, J.; Jansen, T.; Keinert, J.; Serrero, G.; Yue, B.; Stohr, R.; Greither, T.; et al. Combined miR-486 and GP88 (Progranulin) Serum Levels Are Suggested as Supportive Biomarkers for Therapy Decision in Elderly Prostate Cancer Patients. Life 2022, 12, 732. [Google Scholar] [CrossRef]
- Abdulrahman, A.; Eckstein, M.; Jung, R.; Guzman, J.; Weigelt, K.; Serrero, G.; Yue, B.; Geppert, C.; Stohr, R.; Hartmann, A.; et al. Expression of GP88 (Progranulin) Protein Is an Independent Prognostic Factor in Prostate Cancer Patients. Cancers 2019, 11, 2029. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Antonio, R.; Cancino, C.; Arechavaleta-Velasco, F.; Andrade, A.; Barron, L.; Estrada, I.; Fernandez, R.L.; Olguin, V.; Ruiz, S.; Imani, F.; et al. Expression of progranulin (Acrogranin/PCDGF/Granulin-Epithelin Precursor) in benign and malignant ovarian tumors and activation of MAPK signaling in ovarian cancer cell line. Cancer Investig. 2010, 28, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Han, J.J.; Yu, M.; Houston, N.; Steinberg, S.M.; Kohn, E.C. Progranulin is a potential prognostic biomarker in advanced epithelial ovarian cancers. Gynecol. Oncol. 2011, 120, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, A.M.; Maurer, M.J.; Goergen, K.M.; Kalli, K.R.; Erskine, C.L.; Behrens, M.D.; Knutson, K.L.; Block, M.S. Utility of progranulin and serum leukocyte protease inhibitor as diagnostic and prognostic biomarkers in ovarian cancer. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1730–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, D.H.; Do, I.G.; Oh, S.; Lee, Y.G.; Kim, K.; Sohn, J.H.; Park, S.K.; Yang, H.J.; Jung, Y.S.; Park, D.I.; et al. Prognostic Value of Progranulin in Patients with Colorectal Cancer Treated with Curative Resection. Pathol. Oncol. Res. 2020, 26, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Lovat, F.; Bitto, A.; Xu, S.Q.; Fassan, M.; Goldoni, S.; Metalli, D.; Wubah, V.; McCue, P.; Serrero, G.; Gomella, L.G.; et al. Proepithelin is an autocrine growth factor for bladder cancer. Carcinogenesis 2009, 30, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Selmy, M.A.; Ibrahim, G.H.; El Serafi, T.I.; Ghobeish, A.A. Evaluation of urinary proepithelin as a potential biomarker for bladder cancer detection and prognosis in Egyptian patients. Cancer Biomark. 2010, 7, 163–170. [Google Scholar] [CrossRef]
- Soukup, V.; Kalousova, M.; Capoun, O.; Sobotka, R.; Breyl, Z.; Pesl, M.; Zima, T.; Hanus, T. Panel of Urinary Diagnostic Markers for Non-Invasive Detection of Primary and Recurrent Urothelial Urinary Bladder Carcinoma. Urol. Int. 2015, 95, 56–64. [Google Scholar] [CrossRef]
- Eckstein, M.; Lieb, V.; Jung, R.; Sikic, D.; Weigelt, K.; Stöhr, R.; Geppert, C.; Weyerer, V.; Bertz, S.; Serrero, G.; et al. Combination of GP88 Expression in Tumor Cells and Tumor-Infiltrating Immune Cells Is an Independent Prognostic Factor for Bladder Cancer Patients. Cells 2021, 10, 1796. [Google Scholar] [CrossRef]
- Buraschi, S.; Neill, T.; Xu, S.Q.; Palladino, C.; Belfiore, A.; Iozzo, R.V.; Morrione, A. Progranulin/EphA2 axis: A novel oncogenic mechanism in bladder cancer. Matrix Biol. 2020, 93, 10–24. [Google Scholar] [CrossRef]
- Edelman, M.J.; Feliciano, J.; Yue, B.; Bejarano, P.; Ioffe, O.; Reisman, D.; Hawkins, D.; Gai, Q.; Hicks, D.; Serrero, G. GP88 (progranulin): A novel tissue and circulating biomarker for non-small cell lung carcinoma. Hum. Pathol. 2014, 45, 1893–1899. [Google Scholar] [CrossRef] [Green Version]
- Naumnik, W.; Panek, B.; Ossolinska, M.; Naumnik, B. B Cell-Attracting Chemokine-1 and Progranulin in Bronchoalveolar Lavage Fluid of Patients with Advanced Non-small Cell Lung Cancer: New Prognostic Factors. Adv. Exp. Med. Biol. 2019, 1150, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Takemura, M.; Serrero, G.; Yoshikura, N.; Hayashi, Y.; Saito, K.; Inuzuka, T. Higher levels of progranulin in cerebrospinal fluid of patients with lymphoma and carcinoma with CNS metastasis. J. Neurooncol. 2018, 137, 455–462. [Google Scholar] [CrossRef]
- Greither, T.; Steiner, T.; Bache, M.; Serrero, G.; Otto, S.; Taubert, H.; Eckert, A.W.; Kappler, M. GP88/PGRN Serum Levels Are Associated with Prognosis for Oral Squamous Cell Carcinoma Patients. Biology 2021, 10, 400. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Do, I.G.; Kim, K.; Sohn, J.H.; Kim, H.J.; Jeon, W.K.; Lee, S.R.; Son, B.H.; Shin, J.H.; Nam, H.; et al. Progranulin as a predictive factor of response to chemotherapy in advanced biliary tract carcinoma. Cancer Chemother. Pharmacol. 2016, 78, 1085–1092. [Google Scholar] [CrossRef]
- Do, I.G.; Jung, K.U.; Koo, D.H.; Lee, Y.G.; Oh, S.; Kim, K.; Kim, D.H.; Sohn, J.H.; Son, B.H.; Lee, S.R.; et al. Clinicopathological characteristics and outcomes of gastrointestinal stromal tumors with high progranulin expression. PLoS ONE 2021, 16, e0245153. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Park, S.E.; Yun, J.S.; Park, C.Y. Serum Adiponectin and Progranulin Level in Patients with Benign Thyroid Nodule or Papillary Thyroid Cancer. Endocrinol. Metab. (Seoul) 2020, 35, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Tangkeangsirisin, W.; Hayashi, J.; Serrero, G. PC cell-derived growth factor mediates tamoxifen resistance and promotes tumor growth of human breast cancer cells. Cancer Res. 2004, 64, 1737–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrhale, T.; Brodie, A.; Sabnis, G.; Macedo, L.; Tian, C.; Yue, B.; Serrero, G. GP88 (PC-Cell Derived Growth Factor, progranulin) stimulates proliferation and confers letrozole resistance to aromatase overexpressing breast cancer cells. BMC Cancer 2011, 11, 231. [Google Scholar] [CrossRef] [Green Version]
- Pizarro, G.O.; Zhou, X.C.; Koch, A.; Gharib, M.; Raval, S.; Bible, K.; Jones, M.B. Prosurvival function of the granulin-epithelin precursor is important in tumor progression and chemoresponse. Int. J. Cancer 2007, 120, 2339–2343. [Google Scholar] [CrossRef]
- Bose, D.; Zimmerman, L.J.; Pierobon, M.; Petricoin, E.; Tozzi, F.; Parikh, A.; Fan, F.; Dallas, N.; Xia, L.; Gaur, P.; et al. Chemoresistant colorectal cancer cells and cancer stem cells mediate growth and survival of bystander cells. Br. J. Cancer 2011, 105, 1759–1767. [Google Scholar] [CrossRef] [Green Version]
- Wong, N.C.; Cheung, P.F.; Yip, C.W.; Chan, K.F.; Ng, I.O.; Fan, S.T.; Cheung, S.T. Antibody against granulin-epithelin precursor sensitizes hepatocellular carcinoma to chemotherapeutic agents. Mol. Cancer Ther. 2014, 13, 3001–3012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buraschi, S.; Xu, S.Q.; Stefanello, M.; Moskalev, I.; Morcavallo, A.; Genua, M.; Tanimoto, R.; Birbe, R.; Peiper, S.C.; Gomella, L.G.; et al. Suppression of progranulin expression inhibits bladder cancer growth and sensitizes cancer cells to cisplatin. Oncotarget 2016, 7, 39980–39995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, S.T.; Cheung, P.F.; Cheng, C.K.; Wong, N.C.; Fan, S.T. Granulin-epithelin precursor and ATP-dependent binding cassette (ABC)B5 regulate liver cancer cell chemoresistance. Gastroenterology 2011, 140, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.C.; Hsu, J.H.; Lai, L.C.; Tsai, M.H.; Chuang, E.Y. MicroRNA-107 enhances radiosensitivity by suppressing granulin in PC-3 prostate cancer cells. Sci. Rep. 2020, 10, 14584. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Buraschi, S.; Tanimoto, R.; Stefanello, M.; Belfiore, A.; Iozzo, R.V.; Morrione, A. Analysis of Progranulin-Mediated Akt and MAPK Activation. Methods Mol. Biol. 2018, 1806, 121–130. [Google Scholar] [PubMed]
- Tanimoto, R.; Morcavallo, A.; Terracciano, M.; Xu, S.Q.; Stefanello, M.; Buraschi, S.; Lu, K.G.; Bagley, D.H.; Gomella, L.G.; Scotlandi, K.; et al. Sortilin regulates progranulin action in castration-resistant prostate cancer cells. Endocrinology 2015, 156, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Zheng, L.; Zhang, W.; Feng, T.; Liu, J.; Wang, X.; Yu, Y.; Qi, M.; Zhao, W.; Yu, X.; et al. Growth factor progranulin contributes to cervical cancer cell proliferation and transformation in vivo and in vitro. Gynecol. Oncol. 2014, 134, 364–371. [Google Scholar] [CrossRef]
- Feng, T.; Zheng, L.; Liu, F.; Xu, X.; Mao, S.; Wang, X.; Liu, J.; Lu, Y.; Zhao, W.; Yu, X.; et al. Growth factor progranulin promotes tumorigenesis of cervical cancer via PI3K/Akt/mTOR signaling pathway. Oncotarget 2016, 7, 58381. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Li, R.; Wang, H.; Wang, J.; Han, L.; Pan, L.; Li, X.; Kong, Q.; Wang, G.; Su, X. Clinical implications of progranulin in gastric cancer and its regulation via a positive feedback loop involving AKT and ERK signaling pathways. Mol. Med. Rep. 2017, 16, 9685–9691. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.C.; Ip, Y.C.; Cheung, S.T.; Lee, Y.T.; Chan, K.F.; Wong, S.Y.; Fan, S.T. Granulin-epithelin precursor as a therapeutic target for hepatocellular carcinoma. Hepatology 2008, 47, 1524–1532. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, W.; Yang, F.; Feng, T.; Zhou, M.; Yu, Y.; Yu, X.; Zhao, W.; Yi, F.; Tang, W.; et al. Interleukin-6-stimulated progranulin expression contributes to the malignancy of hepatocellular carcinoma cells by activating mTOR signaling. Sci. Rep. 2016, 6, 21260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Bie, M.; Wang, X.; Fan, M.; Chen, B.; Shi, Q.; Jiang, Y. PGRN exacerbates the progression of non-small cell lung cancer via PI3K/AKT/Bcl-2 antiapoptotic signaling. Genes Dis. 2022, 9, 1650–1661. [Google Scholar] [CrossRef]
- Dong, D.; Zhang, W.; Xiao, W.; Wu, Q.; Cao, Y.; Gao, X.; Huang, L.; Wang, Y.; Chen, J.; Wang, W.; et al. A GRN Autocrine-Dependent FAM135B/AKT/mTOR Feedforward Loop Promotes Esophageal Squamous Cell Carcinoma Progression. Cancer Res. 2021, 81, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Frampton, G.; Invernizzi, P.; Bernuzzi, F.; Pae, H.Y.; Quinn, M.; Horvat, D.; Galindo, C.; Huang, L.; McMillin, M.; Cooper, B.; et al. Interleukin-6-driven progranulin expression increases cholangiocarcinoma growth by an Akt-dependent mechanism. Gut 2012, 61, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Li, E.; Luo, L.; Zhao, S.; Liu, L.; Wang, J.; Kang, R.; Luo, J. A PSCA/PGRN-NF-kappaB-Integrin-alpha4 Axis Promotes Prostate Cancer Cell Adhesion to Bone Marrow Endothelium and Enhances Metastatic Potential. Mol. Cancer Res. 2020, 18, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Laudisi, F.; Cherubini, F.; Di Grazia, A.; Dinallo, V.; Di Fusco, D.; Franze, E.; Ortenzi, A.; Salvatori, I.; Scaricamazza, S.; Monteleone, I.; et al. Progranulin sustains STAT3 hyper-activation and oncogenic function in colorectal cancer cells. Mol. Oncol. 2019, 13, 2142–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanimoto, R.; Lu, K.G.; Xu, S.Q.; Buraschi, S.; Belfiore, A.; Iozzo, R.V.; Morrione, A. Mechanisms of Progranulin Action and Regulation in Genitourinary Cancers. Front. Endocrinol. (Lausanne) 2016, 7, 100. [Google Scholar] [CrossRef] [Green Version]
- Tanimoto, R.; Palladino, C.; Xu, S.Q.; Buraschi, S.; Neill, T.; Gomella, L.G.; Peiper, S.C.; Belfiore, A.; Iozzo, R.V.; Morrione, A. The perlecan-interacting growth factor progranulin regulates ubiquitination, sorting, and lysosomal degradation of sortilin. Matrix Biol. 2017, 64, 27–39. [Google Scholar] [CrossRef]
- Kullander, K.; Klein, R. Mechanisms and functions of Eph and ephrin signaling. Nat. Rev. Mol. Cell Biol. 2002, 3, 475–486. [Google Scholar] [CrossRef]
- Zhang, C.; Smalley, I.; Emmons, M.F.; Sharma, R.; Izumi, V.; Messina, J.; Koomen, J.M.; Pasquale, E.B.; Forsyth, P.A.; Smalley, K.S.M. Noncanonical EphA2 Signaling Is a Driver of Tumor-Endothelial Cell Interactions and Metastatic Dissemination in BRAF Inhibitor—Resistant Melanoma. J. Investig. Dermatol. 2021, 141, 840–851.e844. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yamada, N.; Tanaka, T.; Hori, T.; Yokoyama, S.; Hayakawa, Y.; Yano, S.; Fukuoka, J.; Koizumi, K.; Saiki, I.; et al. Crucial roles of RSK in cell motility by catalysing serine phosphorylation of EphA2. Nat. Commun. 2015, 6, 7679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, J.P.; Halford, M.M.; Stacker, S.A. The biochemistry, signalling and disease relevance of RYK and other WNT-binding receptor tyrosine kinases. Growth Factors 2018, 36, 15–40. [Google Scholar] [CrossRef] [PubMed]
- Halford, M.M.; Armes, J.; Buchert, M.; Meskenaite, V.; Grail, D.; Hibbs, M.L.; Wilks, A.F.; Farlie, P.G.; Newgreen, D.F.; Hovens, C.M.; et al. Ryk-deficient mice exhibit craniofacial defects associated with perturbed Eph receptor crosstalk. Nat. Genet. 2000, 25, 414–418. [Google Scholar] [CrossRef]
- Halford, M.M.; Stacker, S.A. Revelations of the RYK receptor. Bioessays 2001, 23, 34–45. [Google Scholar] [CrossRef]
- Adamo, A.; Fiore, D.; De Martino, F.; Roscigno, G.; Affinito, A.; Donnarumma, E.; Puoti, I.; Ricci Vitiani, L.; Pallini, R.; Quintavalle, C.; et al. RYK promotes the stemness of glioblastoma cells via the WNT/ β-catenin pathway. Oncotarget 2017, 8, 13476–13487. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Zavala, M.; Riveros-Magana, A.R.; Garcia-Castro, B.; Barrera-Chairez, E.; Rubio-Jurado, B.; Garces-Ruiz, O.M.; Ramos-Solano, M.; Aguilar-Lemarroy, A.; Jave-Suarez, L.F. WNT receptors profile expression in mature blood cells and immature leukemic cells: RYK emerges as a hallmark receptor of acute leukemia. Eur. J. Haematol. 2016, 97, 155–165. [Google Scholar] [CrossRef]
- Rosen, E.Y.; Wexler, E.M.; Versano, R.; Coppola, G.; Gao, F.; Winden, K.D.; Oldham, M.C.; Martens, L.H.; Zhou, P.; Farese, R.V., Jr.; et al. Functional genomic analyses identify pathways dysregulated by progranulin deficiency, implicating Wnt signaling. Neuron 2011, 71, 1030–1042. [Google Scholar] [CrossRef] [Green Version]
- Alquézar, C.; de la Encarnación, A.; Moreno, F.; López de Munain, A.; Martín-Requero, Á. Progranulin deficiency induces overactivation of WNT5A expression via TNF-α/NF-κB pathway in peripheral cells from frontotemporal dementia-linked granulin mutation carriers. J. Psychiatry Neurosci. 2016, 41, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Alquézar, C.; Esteras, N.; de la Encarnación, A.; Alzualde, A.; Moreno, F.; López de Munain, A.; Martín-Requero, A. PGRN haploinsufficiency increased Wnt5a signaling in peripheral cells from frontotemporal lobar degeneration-progranulin mutation carriers. Neurobiol. Aging 2014, 35, 886–898. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.P.; Tian, Q.Y.; Liu, B.; Cuellar, J.; Richbourgh, B.; Jia, T.H.; Liu, C.J. Progranulin knockout accelerates intervertebral disc degeneration in aging mice. Sci. Rep. 2015, 5, 9102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korade, Z.; Mirnics, K. Wnt signaling as a potential therapeutic target for frontotemporal dementia. Neuron 2011, 71, 955–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Progranulin Autocrine Function on Tumor Cells | |
| Cell proliferation | Colorectal cancer, lung carcinoma, cervical cancer, prostate carcinoma, adrenal carcinoma, laryngeal carcinoma, breast carcinoma. |
| Cell migration and invasion | Breast cancer, colorectal cancer, bladder cancer, prostate carcinoma, adrenal carcinoma, hepatocellular carcinoma, ovarian carcinoma, mesothelioma. |
| CSC maintenance | Hepatocellular carcinoma, breast carcinoma, glioblastoma. |
| Progranulin Modulation of the Tumor Microenvironment | |
| Tumor angiogenesis and lymphangiogenesis | Colorectal cancer, breast cancer, mesothelioma, esophageal squamous cell carcinoma. |
| Tumor immune evasion | Hepatocellular carcinoma, metastatic pancreatic cancer, pancreatic ductal carcinoma, breast carcinoma. |
| Stimulation of fibroblasts and myofibroblasts function | Breast carcinoma, pancreatic ductal adenocarcinoma, colorectal carcinoma. |
| Progranulin Axis as a Biomarker in Cancer | |
| Diagnostic and/or prognostic and/or predictive marker | Breast carcinoma, prostate carcinoma, ovarian epithelial cancers, colorectal carcinoma, bladder cancer, non-small cell lung carcinoma, astrocytoma, glioblastoma, oral squamous cell carcinomas, biliary tract carcinoma, gastrointestinal tumors, papillary thyroid cancer. |
| Progranulin and Resistance to Anticancer Therapies | |
| Chemotherapy | Breast carcinoma, ovarian, colorectal, and hepatocellular carcinomas, glioblastoma, bladder cancer. |
| Radiation therapy | Prostate cancer. |
| Progranulin as a Therapeutic Target in Cancer | |
| Progranulin inhibition via genetic depletion or neutralizing antibodies | Breast carcinoma, ovarian cancer, hepatocellular carcinoma, bladder cancer. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventura, E.; Ducci, G.; Benot Dominguez, R.; Ruggiero, V.; Belfiore, A.; Sacco, E.; Vanoni, M.; Iozzo, R.V.; Giordano, A.; Morrione, A. Progranulin Oncogenic Network in Solid Tumors. Cancers 2023, 15, 1706. https://doi.org/10.3390/cancers15061706
Ventura E, Ducci G, Benot Dominguez R, Ruggiero V, Belfiore A, Sacco E, Vanoni M, Iozzo RV, Giordano A, Morrione A. Progranulin Oncogenic Network in Solid Tumors. Cancers. 2023; 15(6):1706. https://doi.org/10.3390/cancers15061706
Chicago/Turabian StyleVentura, Elisa, Giacomo Ducci, Reyes Benot Dominguez, Valentina Ruggiero, Antonino Belfiore, Elena Sacco, Marco Vanoni, Renato V. Iozzo, Antonio Giordano, and Andrea Morrione. 2023. "Progranulin Oncogenic Network in Solid Tumors" Cancers 15, no. 6: 1706. https://doi.org/10.3390/cancers15061706