Identification of the Invasive Form of Corbicula Clams in Ireland

1
Centre for Environmental Research Innovation and Sustainability (CERIS), Institute of Technology Sligo, Ash Lane, Sligo, F91 YW50, Ireland
2
Monaghan County Council, The Glen, Monaghan Town, Co. Monaghan, H18 YT50, Ireland
3
Department of Biology, Laboratory of Evolutionary Genetics and Ecology (LEGE), University of Namur, Namur, 5000, Belgium
4
Marine Research Institute, Klaipėda University, University Avenue 17, Klaipėda, LT-92294, Lithuania
*
Author to whom correspondence should be addressed.
Water 2019, 11(8), 1652; https://doi.org/10.3390/w11081652
Submission received: 30 June 2019
/
Revised: 31 July 2019
/
Accepted: 5 August 2019
/
Published: 10 August 2019
(This article belongs to the Special Issue Effects of Species Introduction on Aquatic Communities)
Abstract
:The basket clam genus, Corbicula, commonly known as the Asian clam, has become one of the most internationally high-profile and widespread aquatic invasive species. This genus is now considered to comprise a polymorphic species complex. The international invasion of Corbicula is characterised by four lineages, each fixed for one morphotype, genotype and haplotype combination: the American form (A) and European round form (R), the American form (C) and European saddle from (S), American form B, form round light colour (Rlc) and an intermediate between forms R and S known as Int. We investigated the genetic and morphometric makeup of each Irish population in order to establish which invasive lineages were present so as to identify the number of introductions to Ireland. A combination of morphometric, mitochondrial cytochrome oxidase subunit I (mtCOI) gene analysis and microsatellite markers were used to determine the invasive form at each Irish site. All Irish Corbicula samples conformed morphometrically to the invasive form A/R. All mtCOI sequences retrieved for 25 Irish individuals were identical to the international A/R form, while microsatellite markers again showed a common clustering with the international A/R forms of Corbicula. The combined approach of morphometries, total genomic DNA and microsatellite markers indicate only one form of Corbicula invaded Ireland; the international A/R form.
1. Introduction
Corbicula is a freshwater bivalve genus, commonly known as basket clams. Corbicula clams are native to Asia, Australia, Middle East and Africa, and have become invasive in Europe and the Americas [1,2,3,4]. Interestingly, invasive Corbicula clams, as well as most native lineages, reproduce through a peculiar and rare asexual reproductive mode, i.e., androgenesis or strict paternal inheritance. Androgenesis in Corbicula occurs through the expulsion of maternal nuclear chromosomes after obligatory fertilization of an oocyte by an unreduced biflagellate androgenetic spermatozoa [5,6,7].
In North America, the first record of an invasive lineage of Corbicula was made in 1924 in British Columbia [8], with subsequent spread through many parts of the continent (reviewed in [9]). In addition, Corbicula has spread across much of Central and South America during the 1970s and 1980s [3,8,9,10,11]. The first records of Corbicula in Europe were in 1980 from the River Dordogne in France and the Tagus estuary in Portugal [1]. Subsequently, Corbicula has become widespread through much of Western Europe [12] and England [13].
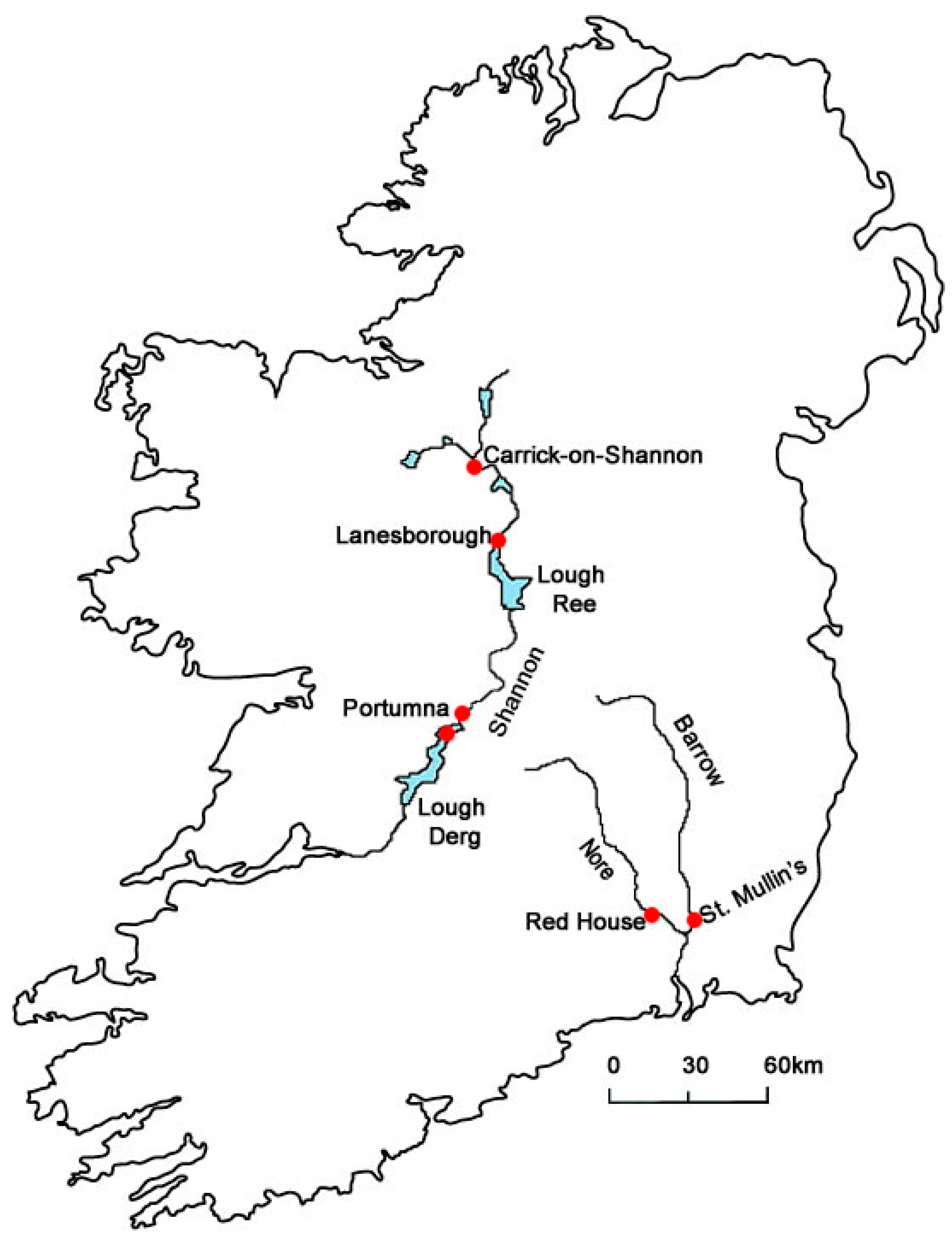
In Ireland, Corbicula clams were first detected in the River Barrow in 2010 [14], where an extremely high density of 17,872 individuals/m2 recorded at one sampling site in 2012 [15]. A subsequent investigation revealed its presence in the River Nore [16], which shares a common estuary with the River Barrow. Corbicula was also discovered in the Shannon system, Ireland’s major navigable waterway, at Carrick-on-Shannon [17], in Lough Derg and in the Shannon River section above Portumna upstream of Lough Derg [18] and at Lanesborough, upstream of Lough Ree (R. Sheehan, Pers Obs) (Figure 1).
Invasive lineages of Corbicula clams have previously been described as belonging to two species, Corbicula fluminea and Corbicula fluminalis but are now assigned to forms [11] based on molecular markers because of the confused taxonomy of the genus and invasive lineages. A morphological taxonomic identification of Corbicula lineages has proved problematic as a wide range of shell morphology exists across invaded countries and within their native Asian range [19,20,21,22,23,24]. The use of mitochondrial analysis and the development of microsatellite markers for invasive Corbicula lineages in Europe, has increased our understanding of both invasive and native populations worldwide [11,24].
Internationally only four lineages of Corbicula have become invasive, with each being fixed for one morphotype, genotype and haplotype combination [11]: form A/R, form C/S, form B and form Rlc [11]. Form A/R is found in North and South America as well as in Europe, being the most widely distributed invasive lineage [8,11,21,25]. This form’s mtCOI haplotype (FW5) has also been recorded in C. leana individuals, an androgenetic Japanese lineage [11]. Form R refers to the strict European populations, whereas form A strictly describes the American forms, but both have the same COI haplotype and microsatellite genotype. Form C/S (haplotype FW17) has been recorded both in South America (form C) and in Europe (form S) and appears related to C. fluminalis [11,21]. Form B, showing the haplotype FW1, is found in North and South America [21]. Finally, form Rlc is found only in Europe, where its distribution appears restricted to the Rhone basin in France [23,26]. Form Rlc’s mitochondrial COI haplotype FW4 is closely related to that of Form B, as they differ only at one nucleotide site but are distinct at the microsatellite genotype [11].
Within invasive populations of Corbicula, further genetic and morphometric complexities can arise from these cryptic hybridizations between sympatric, distinct androgenetic lineages (reviewed in [27]. Indeed, when androgenetic reproduction occurs between two different lineages, the offspring are associated with the nuclear genome and thus, the morphology of the paternal lineage combined to the mitochondrial genome of a distinct maternal lineage, as mitochondria are maternally inherited. This association within one individual of the nuclear genetic information of one lineage and the mitochondrial genetic information of another is called a cytonuclear mismatch [28] and leads to cryptic hybrids. From time to time, nuclear chromosomes can be inadvertently retained within the zygote [6] leading to hybrid individuals with intermediate phenotypes between distinct forms. Such cytonuclear mismatch and hybrid individuals have been widely documented in the Corbicula invasive range [11,19,21,23,28,29]. A recent form D was described in North America [30] but it could be a hybrid between form A/R and form B, as the morphotype is intermediate between these two forms, the COI is from form A/R [23] and the 28S is found in form B [28], as well as in Asia [31]. Moreover, the environment can influence the phenotype, thus complicating the determination of an invasive form when it is based solely on morphotype. For example, differing Corbicula morphotypes in two estuaries in Portugal, possess the same haplotype FW5; their morphological differences are likely due to differing environmental factors in each estuary [22].
The phenomena of cytonuclear mismatches and cryptic hybrids in Corbicula, highlight the importance of combining molecular genetic methods (mitochondrial and nuclear markers) with morphological identification as a dual approach to accurately characterize invasive populations [23].
The pathways and mode by which Corbicula was introduced and has subsequently spread within Ireland are poorly understood [32], with recreational water activity being one possible explanation [33]. It is unclear whether only one discrete introduction occurred, with populations at other sites resulting from secondary spread or alternatively, if a number of separate introductions took place from outside Ireland to each site. Determining distinct morphotypes, genotypes and haplotypes in the various Irish Corbicula populations would support a hypothesis that a number of discrete introductions occurred. The high level of phenotypic plasticity present within the invasive forms of Corbicula [22] and the novel reproductive strategies leading to possible cytonuclear mismatches and cryptic hybrids [23] mean any one method on its own from morphology, mitochondrial and microsatellites is likely to provide an incorrect identification of the invasive from tested. The cross-lineage genetic mixing and recent spread of androgenetic Corbicula lineages [27] and the presence of mismatches between mitochondrial and nuclear markers [11,23,28] further demonstrates the need for a multimethod approach.
Aims
This study aimed to demonstrate the effectiveness of combined morphological and genetic approaches (both mitochondrial and nuclear markers) in resolving invasive bivalve identification. The research also assessed the methodology as an invasion-source tool by using Corbicula specimens from separate invaded sites in Ireland and Belgium.
2. Materials and Methods
2.1. Specimen Collection
Five sites in Ireland with known populations at the time this study was conducted, were sampled to collect individuals for genetic and morphological study (Figure 1; Table 1). One site in Belgium, in the Meuse River at Petit Lanaye (Table 1) was sampled on the 23/10/2013 to collect individuals solely for morphological comparison with individuals from the Irish sites.
A range of standard sampling methods were used to collect Corbicula specimens, depending on water depth and site location, including SCUBA diving, benthic dredge, grab sampler and kick-net. Samples in Ireland were collected from five sites between 2011 and 2013; the Shannon River at Carrick-on-Shannon, Lanesborough, Portumna and Lough Derg, as well as the River Barrow, the River Nore and the Meuse River (Table 1). The Shannon River basin sites were collected from a range of river and lake settings with the St. Mullin’s and Red House sites [16] being riverine.
2.2. Morphological Analysis
A morphological examination of each individual from Ireland (N = 84) and Belgium (N = 51) was carried out, separating them into the morphotypes described in Europe, R, S, Rlc and also, an intermediate morphotype (Int) [19], in order to determine the form of Corbicula according to the morphotype descriptions in [21] and [26]. Form R, (round form) has a shell with well pronounced concentric ridges, is round and broad and generally attains a larger size than form S. Its internal colour is white but may contain pallid purple markings [26]. Form S, (saddle form) again has concentric ridges but these are less raised then in the R form and is narrower and proportionately longer. The internal shell colour is a deep purple throughout. Form Rlc (light colour R from) is superficially similar to the R form in shape, but with a lighter surface shell colour and an off-white to yellow internal colour [26]. Form Int is similar in shape to the R form but with finer less pronounced ridges on the shell [19].
All specimens selected from Ireland and the River Meuse were measured to the nearest mm for shell height (H), length (L) and width (W) using a pair of digital calipers to determine individuals of form R, S and Intermediate (Int), Rlc was not found (Table 2). The ratio between each measurement was calculated for the Irish Corbicula, and compared to individuals of form R, S and Int from the River Meuse. A Principal Component Analysis (PCA) was conducted in RStudio (Version 0.98.501), on the ratios between shell length, height and width, (L/H, L/W, H/W), as described in [21].
2.3. DNA Extraction
Adductor muscle tissue and foot were dissected from each of the Irish specimens for genetic analysis. The samples for the Carrick-on-Shannon, Lanesborough, River Barrow and River Nore sites were preserved in 98% ethanol and stored at ambient temperature. Samples from Lough Derg were initially preserved in methanol and transferred to ethanol after dissection.
Total genomic DNA was extracted from the adductor muscles and/or foot of 50 individual specimens (Table 2), using the DNeasy blood and tissue kit (Qiagen), according to the manufacturer’s protocol. DNA extraction and microsatellite sequencing were carried out at the Laboratory of Evolutionary Genetics and Ecology, (LEGE), University of Namur, Belgium, as previously described in [11,23].
2.4. Mitochondrial COI Gene Analysis
A fragment of 710 bp of the COI gene was amplified in 25 individuals, with representatives from each Irish location. Polymerase Chain Reaction (PCR) was carried out using the universal primers LCO1490 and HCO2198 [34] following the protocol described in [23]. Amplicons were purified and sequenced with the forward universal primer HCO2198 on an automated ABI 3730XL Genetic Analyzer (Genoscreen, Lille, France). Retrieved sequences were visualized, aligned and edited using BioEdit 7.0.5.3 [35]. Corrected sequences and published sequences of the invasive lineages (FW5—Form A/R: GU721082; FW1—Form B: AF196269; FW4—Form Rlc: GU721084; FW17—Form C/S: GU721083) were added to our dataset and used to construct a haplotype median-joining network using the Network 4.6.1.2. [36].
2.5. Microsatellite Marker Analysis
Ten microsatellite markers [24] (ClA01, ClA02, ClA03, ClB03, ClB11, ClC01, ClC08, ClC12, ClE01, and ClD12) were amplified following the protocol of [24]. Microsatellite markers were read on an ABI 3130XL Genetic Analyzer with GeneScan-500 (LIZ) size standard (Applied Biosystems) and scored using GENEMAPPER (Applied Biosystems).
For each of the 50 individuals analysed, we defined a multilocus genotype (here, the unique combination of alleles for the 10 microsatellite loci). These individuals as well as 47 individuals from the European and American invasive individual’s lineages previously typed (10 individuals from Form A/R, 16 from Form B, 10 from Form Rlc and 11 from Form C/S) were clustered based on their multilocus genotype using a discriminant analysis of principal components (DAPC) [37]. The DAPC analysis was performed using the package adegenet [38] implemented in R version 2.15.2 (R development core team 2008). The number of putative populations was first determined using the k-means clustering algorithm [39] for K = 1 to K = 11; 11 being the number of sampled populations added in the analysis. The appropriate number of clusters was defined using the Bayesian Information Criterion (BIC); the value at which the BIC distribution forms an elbow indicating the best clustering (Supplementary Material S1). The relationships between the BIC-defined clusters were then inferred. Six principal components and one discriminant function (98.9% of the total variance) were retained to represent the majority of the variability contained in the dataset.
3. Results
3.1. Morphology
From the morphological characterization, all Irish Corbicula clams were visually determined as the European form R as described in [19,23] with a round deep shell that may range from externally dark to golden, heavy ridges and a generally white interior (Figure 2).
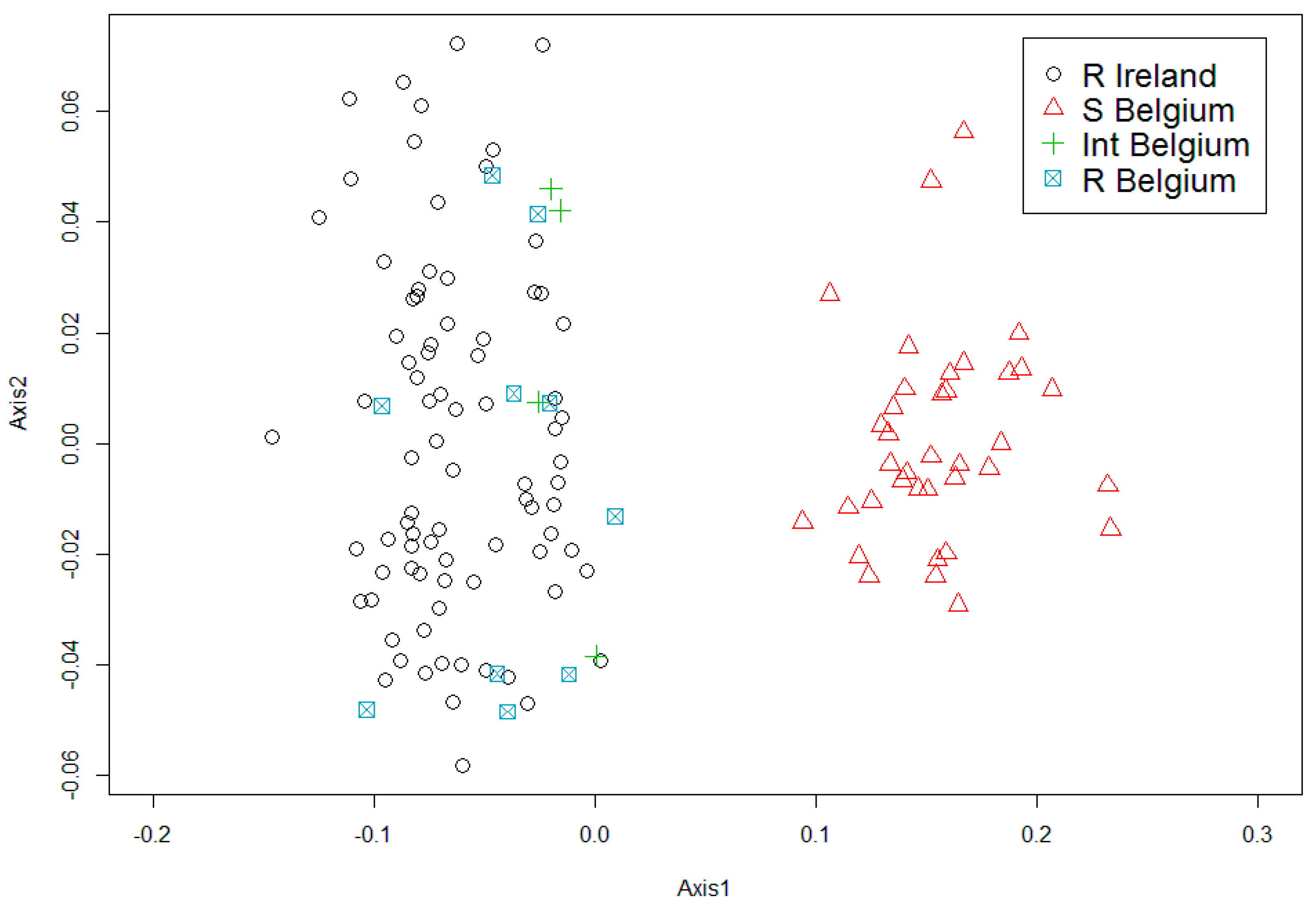
The PCA analysis (Figure 3) of shell height vs length shows the Irish Corbicula grouping together with the Belgium form R and the international intermediate (Int) form. The Int form displayed an intermediate morphology between R and S, with the round body shape of R and the closely spaced shell ridges and purple interior coloring of S. The Irish individuals were different from the Belgian form S. No Irish individuals had a shell height/length ratio consistent with form S.
3.2. COI Sequence
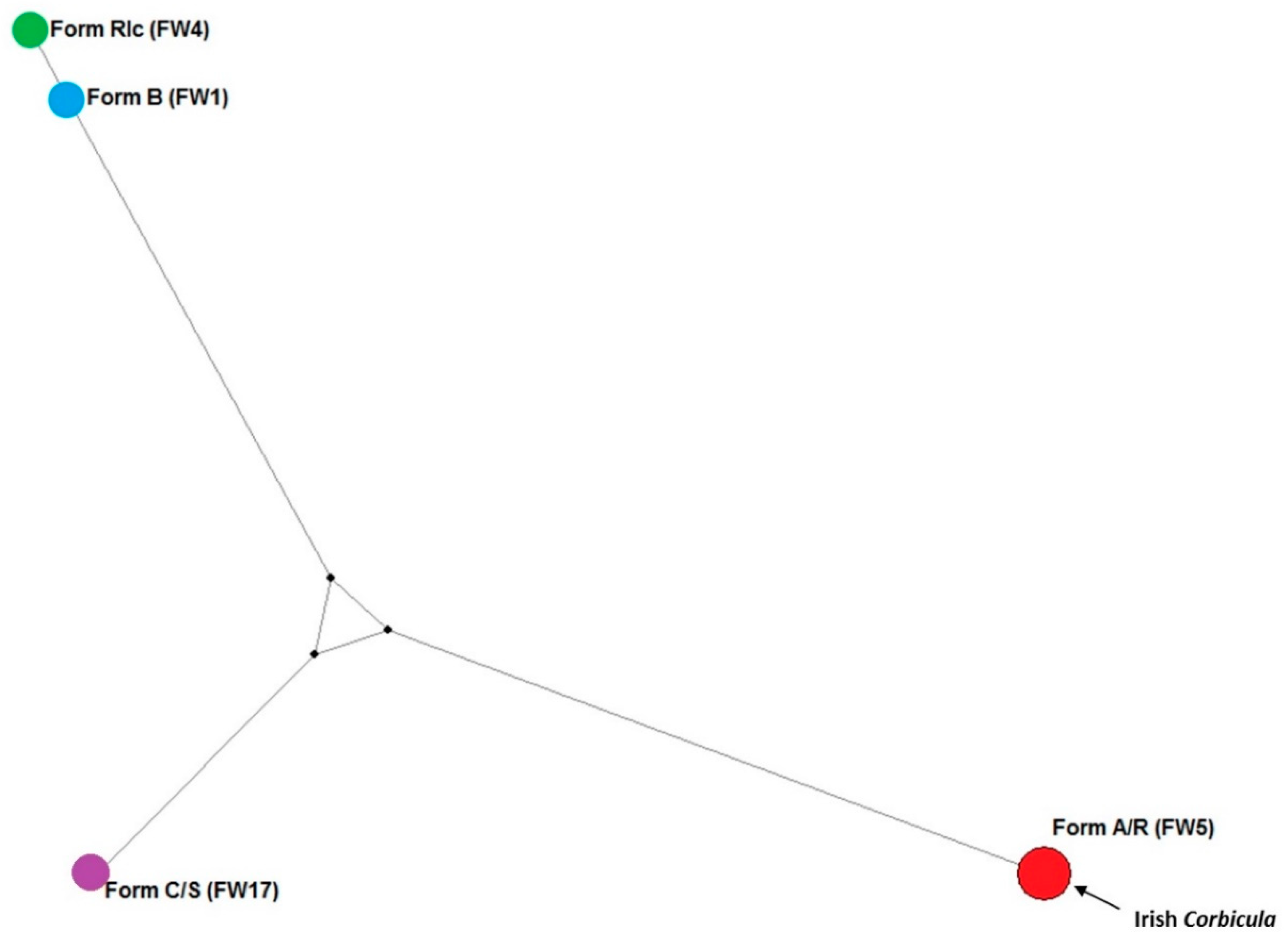
A 710 bp length fragment of the mitochondrial COI was successfully amplified for 24 of the Irish specimens (Table 2). All the COI sequences retrieved were identical to FW5, the mtCOI haplotype of form A/R distributed in Europe and Americas. All individuals sampled from the Irish populations possessed the same haplotype. No haplotypes from other invasive lineages (Figure 4) were detected from the Irish sites.
3.3. Microsatellite Markers
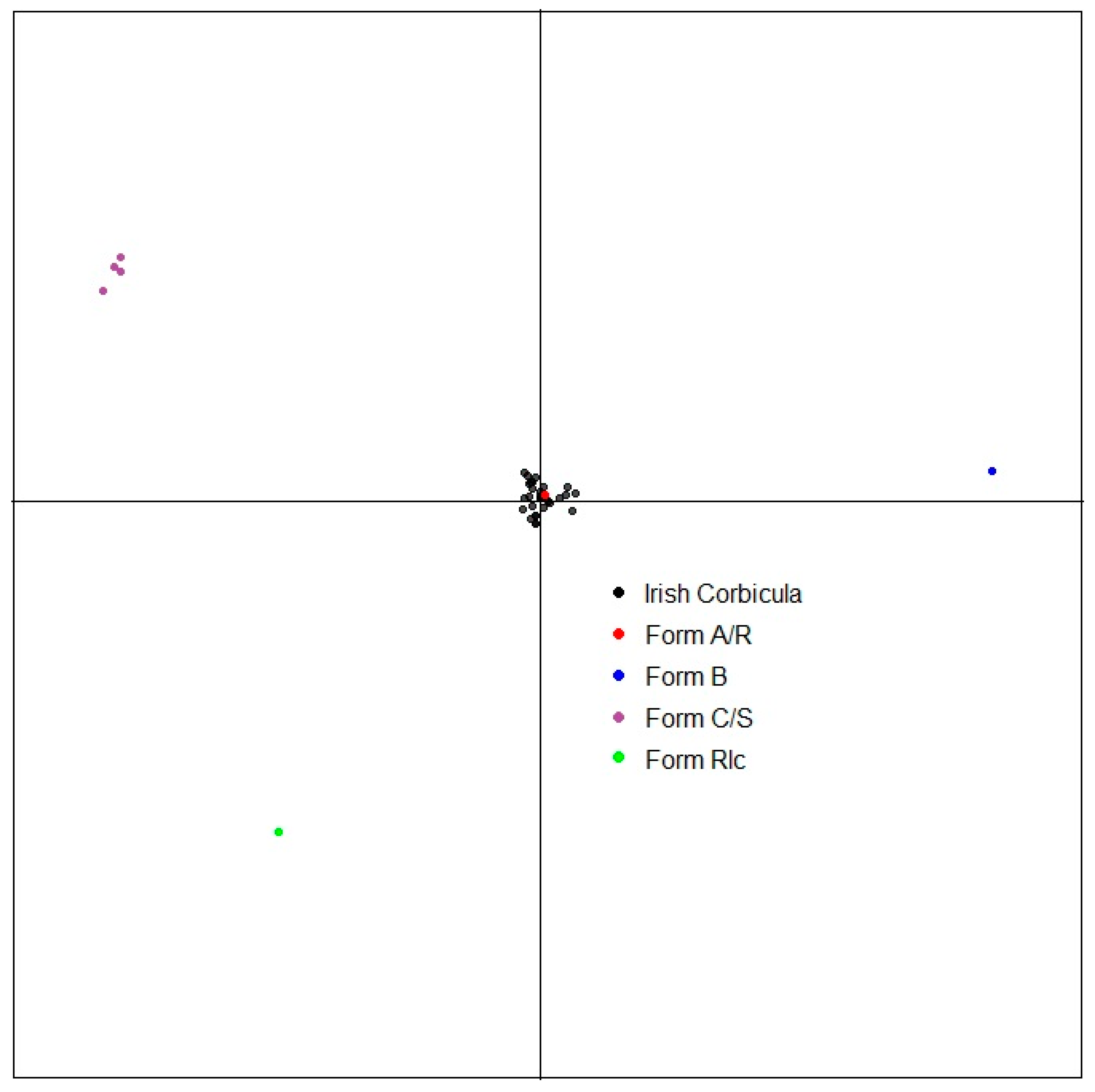
Genetic diversity and genetic relationships in invasive Corbicula clams estimated through a Discriminant Analysis of Principal Components (Figure 5) based on multilocus genotypes from microsatellite marker amplifications show a common clustering between all Irish samples and the American and European invasive form A/R, suggesting that Irish Corbicula belongs to this same lineage.
4. Discussion
Here, we used a multi-method approach, including mitochondrial COI sequencing, microsatellite genotyping and morphological analysis, to identify the invasive Corbicula lineages in Ireland. Such an integrative approach was necessary as the genus Corbicula is characterized by haplotypes capable of displaying highly divergent phenotypes in response to differing environmental conditions or following cross-lineages mixing.
Our results showed that only the invasive form A/R, the most widespread invasive lineage, was present from all samples collected in Irish waters, as determined using a combination of (a) morphological analysis, (b) mtDNA (COI) and (c) microsatellite markers.
No clams with either a narrow and fine-ridged shell or deep purple interior, as corresponding to form S [19,40] were observed. The PCA analysis carried out on the ratio between shell length and shell height (Figure 3) supports the correct classification of these individuals a belonging to form R. Morphologically, all clams sampled from the Irish sites conform to the invasive form A/R as described in [11,21,23,28,29], with a certain variability observed (Figure 3).
All individuals sampled from the Irish sites presented the invasive FW5 mtCOI haplotype of form A/R distributed in Europe and Americas [11,21,23,28,29]. The COI sequence data supports the results from the morphological analysis: no mismatches were observed between mtCOI and morphotype, which may be potentially caused by reproduction through androgenesis between distinct lineages [28].
All Irish sampled individuals showed a common clustering for the microsatellite data with the A/R lineage. This form has proven to be clonal and, thus, shows no genetic diversity. Indeed hundreds of individuals of this lineage, sampled at different locations from across Europe, North and South America, present the exact same multilocus genotype with no genetic diversity [11]. The genetic variability detected within Irish populations is very likely due to the poor amplification of the microsatellites (Supplementary Material S2). Initial storage conditions of some samples in methanol was likely responsible for this. It is also a possible that false positives were detected from the data.
It is unlikely that other forms of Corbicula exist in Ireland as extensive investigations have not revealed clams displaying differing morphological characteristics [14,16,17] (Minchin Pers Ob.) or in subsequentially discovered populations [41,42]. As the level of genetic diversity among the invasive lineages of Corbicula clams (reviewed in [18]) was low, it was not possible to discriminate the origin of the Irish populations. The emerging field of massive parallel sequencing (MPS) to detect single nucleotide polymorphisms (SNPs) [43] could potentially complement the approaches used in our study. Similarly, it is not possible to draw any inferences on the number of discrete introductions that may have occurred [44].
The combined methodology of mitochondrial COI sequencing, microsatellite genotyping and morphological analysis has the potential to pick apart the exact identification of genetically complex species groups for which traditional ecological methods may have overlooked complex relationships such as in Corbicula [27] with its highly invasive clonal A/R form. The ability to discriminate between discreet introductions to a geographic area, and secondary spread is an invaluable tool in prioritising AIS biosecurity resources [45] and can inform horizon scanning [46].
5. Conclusions
The use of a combined morphological and nuclear marker approach in resolving the identity of the invasive Corbicula, demonstrates that only one invasive lineage of Corbicula has invaded Ireland, the most prevalent and widely distributed form being A/R, a form also found across Europe and America. The extremely low genetic diversity found within this invasive lineage makes the determination of differences in population origins difficult as a result, and therefore, the number of discrete introductions of Corbicula to Ireland remains unknown. In order to properly inform management plans and limit impacts to ecosystem services, the identity of invasive Corbicula populations must be resolved. The use of a combined morphological, mitochondrial and nuclear marker approach in gleaning the identity of invasive Corbicula, as demonstrated in this study, provides a useful tool for achieving these goals, with the possibility for extending this approach to other invasive species.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/11/8/1652/s1, supplementary S1, supplementary S2.
Author Contributions
Conceptualization, R.S., E.E. and F.L.; Methodology, R.S., E.E., F.L., K.V.D. and D.M.; Software, R.S. and E.E.; Validation, R.S. and E.E.; Formal analysis, R.S. and E.E.; Investigation, R.S, and E.E.; Resources, F.L. and K.V.D.; Data Curation, R.S., E.E., F.L. and K.V.D.; Writing—Original Draft Preparation, R.S. and E.E.; Writing—Review and Editing, R.S., E.E., F.L., D.M. and K.V.D.; Visualization, R.S. and E.E.; Supervision, F.L. and K.V.D.; Project Administration, R.S., E.E., F.L. and K.V.D.; Funding Acquisition, F.L. and K.V.D.
Funding
This research was funded by President’s bursary award, Institute of Technology, Sligo. Financial support was also provided by the COST Action no. TD1209 ‘Alien species: Linking Information across a European Network (ALIEN Challenge)’ fund. Financial support from Inland Fisheries Ireland (IFI) is also acknowledged.
Acknowledgments
Particular thanks to Alan Cullagh, Declan Cullagh, IFI Clonmel. Fergus Lynch and all of the IFI Drumsna staff. Joe Caffrey and Michael Millane of IFI Citywest. Julie Virgo of UNamur and Helen Moran of Geomara, Clarinbridge, Co. Galway, Ireland.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mouthon, J. Sur la présence en France et au Portugal de Corbicula (Bivalvia, Corbiculidae) originaire d’Asie. Basteria 1981, 45, 109–116. [Google Scholar]
- McMahon, R.F. The occurrence and spread of the introduced Asiatic fresh-water clam, Corbicula-fluminea (Müller), in North-America-1924-1982. Nautilus 1982, 96, 134–141. [Google Scholar]
- Kraemer, L.R.; Galloway, M.L. Larval development of Corbicula-fluminea (Muller) (Bivalvia, Corbicula Cea)—An appraisal of its heterochrony. Am. Malacol. Bull. 1986, 4, 61–79. [Google Scholar]
- Araujo, R.; Moreno, D.; Ramos, M.A. The Asiatic clam Corbicula fluminea (Müller, 1774)(Bivalvia: Corbiculidae) in Europe. Am. Malacol. Bull. 1993, 10, 39–49. [Google Scholar]
- Komaru, A.; Kawagishi, T.; Konishi, K. Cytological evidence of spontaneous androgenesis in the freshwater clam Corbicula leana Prime. Dev. Genes Evol. 1998, 208, 46–50. [Google Scholar] [CrossRef]
- Komaru, A.; Ookubo, K.; Kiyomoto, M. All meiotic chromosomes and both centrosomes at spindle pole in the zygotes discarded as two polar bodies in clam Corbicula leana: Unusual polar body formation observed by antitubulin immunofluorescence. Dev. Genes Evol. 2000, 210, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Pigneur, L.M.; Hedtke, S.M.; Etoundi, E.; Van Doninck, K. Androgenesis: A review through the study of the selfish shellfish Corbicula spp. Heredity 2012, 108, 581. [Google Scholar] [CrossRef]
- Counts, C.L. Corbicula fluminea (Bivalvia: Sphaeriacea) in British Columbia. Nautilus 1981, 95, 12–13. [Google Scholar]
- Matthews, M.A.; McMahon, R.F. Effects of temperature and temperature acclimation on survival of zebra mussels (Dreissena polymorpha) and Asian clams (Corbicula fluminea) under extreme hypoxia. J. Molluscan Stud. 1999, 65, 317–325. [Google Scholar] [CrossRef]
- Ituarte, C.F. Corbicula and Neo Corbicula (Bivalvia: Corbiculidae) in the Paraná, Uruguay and Río de La Plata Basins. Nautilus 1994, 107, 129–135. [Google Scholar]
- Pigneur, L.; Etoundi, E.; Aldridge, D.C.; Marescaux, J.; Yasuda, N.; Van Doninck, K. Genetic uniformity and long-distance clonal dispersal in the invasive androgenetic Corbicula clams. Mol. Ecol. 2014, 23, 5102–5116. [Google Scholar] [CrossRef] [PubMed]
- Delivering Alien Invasive Species Inventories for Europe (DAISIE). Available online: http://www.europe-aliens.org/ (accessed on 20 May 2007).
- Aldridge, D.C.; Müller, S.J. The Asiatic clam, Corbicula fluminea, in Britain: Current status and potential impacts. J. Conchol. 2001, 37, 177–183. [Google Scholar]
- Sweeney, P. First record of Asian Clam (Corbicula fluminea (Müller, 1774) in Ireland. Irish Nat. J. 2009, 30, 147–148. [Google Scholar]
- Sheehan, R.; Caffrey, J.M.; Millane, M.; McLoone, P.; Moran, H.; Lucy, F. An investigation into the effectiveness of mechanical dredging to remove Corbicula fluminea (Müller, 1774) from test plots in an Irish river system. Manag. Biol. Invasions 2014, 5, 407–418. [Google Scholar] [CrossRef]
- Caffrey, J.M.; Evers, S.; Millane, M.; Moran, H. Current status of Ireland’s newest invasive species-the Asian clam Corbicula fluminea (Mueller, 1774). Aquat. Invasions 2011, 6, 291–299. [Google Scholar] [CrossRef]
- Hayden, B.; Caffrey, J.M. First recording of the Asian Clam (Corbicula fluminea (Müller, 1774)) from the River Shannon, with preliminary notes on population size and size class distribution. Irish Nat. J. 2013, 32, 29–31. [Google Scholar]
- Minchin, D. The distribution of the Asian clam Corbicula fluminea and its potential to spread in Ireland. Manag Biol Invasions 2014, 5, 165–177. [Google Scholar] [CrossRef]
- Pfenninger, M.; Reinhardt, F.; Streit, B. Evidence for cryptic hybridization between different evolutionary lineages of the invasive clam genus Corbicula (Veneroida, Bivalvia). J. Evol. Biol. 2002, 15, 818–829. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, W. Two Corbicula (Corbiculidae: Bivalvia) mitochondrial lineages are widely distributed in Asian freshwater environment. Mol. Phylogenet. Evol. 2003, 29, 529–539. [Google Scholar] [CrossRef]
- Lee, T.; Siripattrawan, S.; Ituarte, C.F.; Foighil, D.O. Invasion of the clonal clams: Corbicula lineages in the New World. Am. Malacol. Bull. 2005, 20, 113–122. [Google Scholar]
- Sousa, R.; Freire, R.; Rufino, M.; Méndez, J.; Gaspar, M.; Antunes, C.; Guilhermino, L. Genetic and shell morphological variability of the invasive bivalve Corbicula fluminea (Müller, 1774) in two Portuguese estuaries. Estuar. Coast. Shelf Sci. 2007, 74, 166–174. [Google Scholar] [CrossRef]
- Pigneur, L.M.; Marescaux, J.; Roland, K.; Etoundi, E.; Descy, J.P.; Van Doninck, K. Phylogeny and androgenesis in the invasive Corbicula clams (Bivalvia, Corbiculidae) in Western Europe. BMC Evol. Biol. 2011, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Pigneur, L.M.; Risterucci, A.M.; Dauchot, N.; Li, X.; Van Doninck, K. Development of novel microsatellite markers to identify the different invasive lineages in the Corbicula complex and to assess androgenesis. Mol. Ecol. Resour. 2011, 11, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Patton, J.C. Morphological and electrophoretic evidence for two species of Corbicula (Bivalvia: Corbiculidae) in North America. Am. Midl. Nat. 1982, 108, 74–80. [Google Scholar] [CrossRef]
- Marescaux, J.; Pigneur, L.M.; Van Doninck, K. New records of Corbicula clams in French rivers. Aquat. Invasions 2010, 5, S35–S39. [Google Scholar] [CrossRef]
- Etoundi, E.; Marescaux, J.; Vastrade, M.; Debortoli, N.; Hedtke, S.M.; Pigneur, L.M.; Virgo, J.; Flot, J.F.; Van Doninck, K. Distinct biogeographic origins of androgenetic Corbicula lineages followed by genetic captures. bioRxiv 2019, 590836. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Stanger-Hall, K.; Baker, R.J.; Hillis, D.M. All-male asexuality: Arigin and maintenance ofandrogenesis in the Asian clam Corbicula. Evolution 2008, 62, 1119–1136. [Google Scholar]
- Park, J.K.; Lee, J.S.; Kim, W. A single mitochondrial lineage is shared by morphologically and allozymatically distinct freshwater Corbicula clones. Mol. Cells 2002, 14, 318–322. [Google Scholar]
- Tiemann, J.S.; Haponski, A.E.; Douglass, S.A.; Lee, T.; Cummings, K.S.; Davis, M.A.; Foighil, D.Ó. First record of a putative novel invasive Corbicula lineage discovered in the Illinois River, Illinois, USA. BioInvasions Rec. 2017, 6, 159–166. [Google Scholar] [CrossRef]
- Komaru, A.; Yamada, M.; Houki, S. Relationship between two androgenetic clam species, Corbicula leana and Corbicula fluminea, inferred from mitochondrial cytochrome b and nuclear 28S rRNA markers. Zoolog. Sci. 2013, 30, 360–366. [Google Scholar] [CrossRef]
- Lucy, F.E.; Karatayev, A.Y.; Burlakova, L.E. Predictions for the spread, population density, and impacts of Corbicula fluminea in Ireland. Aquat. Invasions 2012, 7, 465–474. [Google Scholar] [CrossRef]
- Minchin, D.; Boelens, R. Natural dispersal of the introduced Asian clam Corbicula fluminea (Müller, 1774) (Cyrenidae) within two temperate lakes. Bioinvasions Rec. 2018, 7, 259–268. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Series 1999, 41, 95–98. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Legendre, L.; Legrendre, L. Numerical Ecology. Developments in Environmental Modelling; Elsevier Science & Technology: Amsterdam, The Nederland, 1998; ISBN 1281021598. [Google Scholar]
- Renard, E.; Bachmann, V.; Cariou, M.L.; Moreteau, J.C. Morphological and molecular differentiation of invasive freshwater species of the genus Corbicula (Bivalvia,Corbiculidea) suggest the presence of three taxa in French rivers. Mol. Ecol. 2000, 9, 2009–2016. [Google Scholar] [CrossRef]
- Minchin, D. The Asian Clam (Corbicula fluminea (OF Müller))(Cyrenidae) arrives in the Erne River. Irish Nat. J. 2017, 35, 99–104. [Google Scholar]
- Caffrey, J.M.; Dick, J.T.A.; Lucy, F.E.; Davis, E.; Niven, A.; Coughlan, N.E. First record of the Asian clam Corbicula fluminea (Müller, 1774)(Bivalvia, Cyrenidae) in Northern Ireland. BioInvasions Rec. 2016, 5, 239–244. [Google Scholar] [CrossRef]
- Peñarrubia, L.; Viñas, J.; Sanz, N.; Smith, B.L.; Bremer, J.R.A.; Pla, C.; Vidal, O. SNP identification and validation in two invasive species: Zebra mussel (Dreissena polymorpha) and Asian clam (Corbicula fluminea). Anim. Biodivers. Conserv. 2019, 42, 65–68. [Google Scholar] [CrossRef]
- Schmidlin, S.; Schmera, D.; Ursenbacher, S.; Baur, B. Separate introductions but lack of genetic variability in the invasive clam Corbicula spp. in Swiss lakes. Aquat. Invasions 2012, 7, 73–80. [Google Scholar] [CrossRef]
- Gherardi, F.; Aquiloni, L.; Diéguez-Uribeondo, J.; Tricarico, E. Managing invasive crayfish: Is there a hope? Aquat. Sci. 2011, 73, 185–200. [Google Scholar] [CrossRef]
- González-Moreno, P.; Lazzaro, L.; Vilà, M.; Preda, C.; Adriaens, T.; Bacher, S.; Brundu, G.; Copp, G.H.; Essl, F.; García-Berthou, E. Consistency of impact assessment protocols for non-native species. NeoBiota 2019, 44, 1–25. [Google Scholar] [CrossRef]
Figure 1.
Irish rivers containing Corbicula specimens, previously described as C. fluminea, sampled during this study, River Nore, Barrow and Shannon. Sampling sites Carrick-on-Shannon, Lanesborough, Portumna, Lough Derg, St. Mullin’s and Red House.
Figure 1.
Irish rivers containing Corbicula specimens, previously described as C. fluminea, sampled during this study, River Nore, Barrow and Shannon. Sampling sites Carrick-on-Shannon, Lanesborough, Portumna, Lough Derg, St. Mullin’s and Red House.

Figure 2.
Examples of Corbicula form R from the River Shannon at Lanesborough (juvenile specimen) and Carrick-on-Shannon and the River Barrow at St. Mullin’s. Also shown is an example of Form S from the River Meuse at Maastricht.
Figure 2.
Examples of Corbicula form R from the River Shannon at Lanesborough (juvenile specimen) and Carrick-on-Shannon and the River Barrow at St. Mullin’s. Also shown is an example of Form S from the River Meuse at Maastricht.

Figure 3.
A principal component analysis (PCA) of the ratio between shell measurements for Corbicula form R from all Irish sites and forms R, S and individuals with intermediate morphotypes between form R and S (Int), from the River Meuse in Belgium.
Figure 3.
A principal component analysis (PCA) of the ratio between shell measurements for Corbicula form R from all Irish sites and forms R, S and individuals with intermediate morphotypes between form R and S (Int), from the River Meuse in Belgium.

Figure 4.
Haplotypic diversity and relationships in Invasive Corbicula clams inferred through median-joining Network. The European invasive haplotypes FW1 (Form B), FW4 (Form Rlc), FW17 (Form C/S), FW5 (Form A/R; Irish individuals) are plotted. Branch length was proportional to the number of mutations between the haplotypes and nodes proportional to the haplotype frequency.
Figure 4.
Haplotypic diversity and relationships in Invasive Corbicula clams inferred through median-joining Network. The European invasive haplotypes FW1 (Form B), FW4 (Form Rlc), FW17 (Form C/S), FW5 (Form A/R; Irish individuals) are plotted. Branch length was proportional to the number of mutations between the haplotypes and nodes proportional to the haplotype frequency.

Figure 5.
Genetic diversity and genetic relationships in invasive Corbicula clams estimated through a Discriminant Analysis of Principal Components based on multilocus genotypes. Only axes 1 and 2 are represented. One dot represents one distinct multilocus genotype; individuals showing the exact same genotype are, therefore, pictured under the same dot.
Figure 5.
Genetic diversity and genetic relationships in invasive Corbicula clams estimated through a Discriminant Analysis of Principal Components based on multilocus genotypes. Only axes 1 and 2 are represented. One dot represents one distinct multilocus genotype; individuals showing the exact same genotype are, therefore, pictured under the same dot.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sampling sites and collection methods of Corbicula clams for DNA and morphometric analysis.
Table 1.
Sampling sites and collection methods of Corbicula clams for DNA and morphometric analysis.
| Site | No | Site | Sampling Method | Grid Reference |
|---|---|---|---|---|
| Shannon (Carrick-on-Shannon) | 1 | 1 km above town | Diving | 53.951402, −8.109267 |
| Shannon (Lanesborough) | 2 | Below bridge | Grab + dredge | 53.669457, −7.998963 |
| Shannon (Portumna) | 3 | Below bridge | Grab + dredge | 53.091139, −8.195643 |
| Lough Derg | 4 | Upper lake | Grab + dredge | 52.928985, −8.280803 |
| River Barrow | 5 | St Mullin’s | Diving | 52.487025, −6.926753 |
| River Nore | 6 | Redhouse | Kick-net | 52.469403, −7.044622 |
| River Meuse | 7 | Meuse River, Petit Lanaye, Belgium | Kick-net | 50.810898, −5.692482 |
Table 2.
Numbers of individuals (forms R, S and Intermediate (Int)) used for the morphological (Ireland and Belgium), mtCOI and microsatellites (Ireland only) analyses.
Table 2.
Numbers of individuals (forms R, S and Intermediate (Int)) used for the morphological (Ireland and Belgium), mtCOI and microsatellites (Ireland only) analyses.
| Site | Described Form | Morphology | mtCOI | Microsatellites |
|---|---|---|---|---|
| Carrick-on-Shannon | R | 11 | 5 | 9 |
| Lanesborough | R | 9 | 5 | 9 |
| Lough Derg | R | 39 | 5 | 14 |
| River Barrow | R | 10 | 4 | 9 |
| Portumna | R | 10 | 1 | 2 |
| River Nore | R | 5 | 5 | 5 |
| Meuse River | R | 10 | N/A | N/A |
| S | 37 | N/A | N/A | |
| Int | 4 | N/A | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sheehan, R.; Etoundi, E.; Minchin, D.; Doninck, K.V.; Lucy, F. Identification of the Invasive Form of Corbicula Clams in Ireland. Water 2019, 11, 1652. https://doi.org/10.3390/w11081652
AMA Style
Sheehan R, Etoundi E, Minchin D, Doninck KV, Lucy F. Identification of the Invasive Form of Corbicula Clams in Ireland. Water. 2019; 11(8):1652. https://doi.org/10.3390/w11081652
Chicago/Turabian StyleSheehan, Rory, Emilie Etoundi, Dan Minchin, Karine Van Doninck, and Frances Lucy. 2019. "Identification of the Invasive Form of Corbicula Clams in Ireland" Water 11, no. 8: 1652. https://doi.org/10.3390/w11081652
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.