





Abstract
Recently burned coniferous forests host wildlife communities that respond to variation in burn severity, post-fire habitat structure, and patch configuration. Habitat selection theory predicts that birds inhabiting these variable post-fire landscapes will select nesting locations that confer an adaptive advantage through increased fitness and reproductive success. Understanding the effect of post-fire habitat on avian nesting ecology can provide valuable information to guide restoration and management after wildfire. The Black-backed Woodpecker (Picoides arcticus) is strongly associated with recently burned forests in the western United States, where it is used as an indicator species for the effects of post-fire forest management. Between 2011 and 2018, we located and monitored 118 Black-backed Woodpecker nests in burned forests of northern California. We evaluated the influence of habitat and nest characteristics on nest site selection and daily nest survival. Our results demonstrate a pattern of neutral congruence between habitat selection and fitness. Black-backed Woodpeckers showed strong selection for each of the nest habitat variables that we measured: woodpeckers selected moderately sized trees in areas of high snag density burned at high severity, but also in areas relatively close to low-severity or unburned edges. However, only nest initiation date affected nest survival, with decreased survival in late-season nests. Our results suggest that management actions aimed at maintaining breeding habitat for Black-backed Woodpeckers should prioritize retention and creation of pyrodiverse landscapes that include dense stands of snags (>5 snags per 100 m2) within ~500 m of forest that burned at low severity or remained unburned.
Resumen
Los bosques de coníferas recientemente quemados albergan comunidades de vida silvestre que responden a la intensidad del fuego, a la estructura del hábitat luego del fuego y a la configuración de los parches. La teoría de la selección de hábitat predice que las aves que habitan estos paisajes incendiados diversos seleccionarán aquellas ubicaciones de los nidos que les confieran una ventaja adaptativa a través de un aumento en la adecuación biológica y en el éxito reproductivo. Entender el efecto del hábitat incendiado en la ecología de anidación de las aves puede brindar información valiosa para guiar la restauración y el manejo luego de los fuegos silvestres. Picoides arcticus está fuertemente asociado con bosques recientemente quemados en el oeste de EEUU, donde es usado como una especie indicadora de los efectos del manejo del bosque luego del fuego. Entre 2011 y 2018, localizamos y monitoreamos 118 nidos de P. arcticus en bosques quemados del norte de California. Evaluamos la influencia de las características del hábitat y del nido en la selección del nido y en la supervivencia diaria del nido. Nuestros resultados demuestran un patrón de congruencia neutral entre la selección del hábitat y la adecuación biológica. P. arcticus mostró una fuerte selección de cada una de las variables del hábitat del nido que medimos: los carpinteros seleccionaron árboles de tamaño moderado en áreas de alta densidad de tocones severamente quemados, pero también en áreas relativamente cercanas a bordes con baja severidad o no quemados. Sin embargo, solo la fecha de inicio del nido afectó la supervivencia del nido, con una disminución en la supervivencia de los nidos de fines de estación. Nuestros resultados sugieren que las acciones de manejo destinadas al mantenimiento del hábitat reproductivo para P. arcticus debería priorizar la retención y creación de paisajes pirodiversos que incluyan rodales densos de tocones (>5 tocones por 100 m2) dentro de los ~500 m de bosque quemado con baja severidad o que permaneció no quemado.
INTRODUCTION
Before raising young, birds must make a series of decisions at a variety of spatial scales regarding the location of breeding activities (Hildén 1965, Johnson 1980). These behavioral choices often result in fitness consequences manifested through nest success, such that selected habitats yield increased fitness (Hildén 1965, Martin 1998, Chalfoun and Schmidt 2012). However, empirical studies of nesting ecology do not always demonstrate positive relationships, or congruence, between the factors important for nest site selection and the factors that affect fitness (Chalfoun and Schmidt 2012). For example, incongruent relationships between habitat selection and fitness may arise if settlement cues become decoupled from their historical outcomes, as in ecological traps (Schlaepfer et al. 2002). In a review of studies on avian nesting ecology, Chalfoun and Schmidt (2012) found that over 50% of studies reported some form of neutral congruence, where habitat had an effect on nest site selection without showing a detectible effect on reproductive success.
The high prevalence of neutral congruence in avian breeding biology underscores the importance of considering the relationship between nest site selection and nest success (the probability that at least one nestling fledges) when evaluating management actions that affect bird populations. Effective management will both promote suitable breeding habitat to increase breeding bird abundance and promote the habitat characteristics within selected habitats that increase the probability of nest success (Chalfoun and Martin 2007). Studies that link nest site selection and nest success may also reveal separate effects of habitat—if habitat influences one process but not the other, managers can leverage this information to guide where and how to efficiently invest management resources. This may prove especially valuable in management of burned forests, where management decisions are often time sensitive and may have significant impacts on post-fire bird communities (Noss et al. 2006, Hanson and North 2008).
The Black-backed Woodpecker (Picoides arcticus) is a habitat specialist strongly associated with high densities of standing dead trees (snags) used for foraging and nesting substrates (Hutto 2008, Nappi and Drapeau 2009, Tremblay et al. 2016). Throughout their range in the montane western United States, Black-backed Woodpeckers are predominantly found within recently burned forests, where they forage primarily on the larvae of wood-boring beetles that rapidly colonize dead and dying trees after wildfire (Murphy and Lehnhausen 1998). Previous research in the western United States has demonstrated selection for territories and nest sites in areas with high snag densities created by severe wildfire (Seavy et al. 2012, Tingley et al. 2014), although the species is also found in lower densities in unburned forest (Fogg et al. 2014). Black-backed Woodpecker abundance generally peaks within 5 yr after fire, followed by sharp declines as snags deteriorate (Tingley et al. 2018).
Given the specialized habitat preferences of Black-backed Woodpeckers, habitat selection theory predicts that individuals should select nest locations based on habitat conditions that correlate with increased nest success. Dense snag stands that burned at high severity likely provide increased access to food resources for Black-backed Woodpeckers (Murphy and Lehnhausen 1998, Nappi et al. 2003), and nest success may be higher for nests near foraging habitat. Nest success also may reflect proximity to edge habitats; nests of other woodpecker species located nearer to habitat edges (i.e. ecotones between high- and low-severity burn patches) have shown lower daily survival rates due to higher predation rates (Fisher and Wiebe 2006, Saab et al. 2011). At a finer scale, nest success may respond to microhabitat characteristics such as tree diameter (Wiebe 2001), nest cavity orientation (Hooge et al. 1999), and nest height (Fisher and Wiebe 2006), which influence nest temperature and predation risk.
When neutral congruence exists between the habitat characteristics that affect nest site selection and nest success, we might expect daily nest survival to be primarily driven by a variety of abiotic/temporal variables. For example, nest initiation date may relate to breeding success because of resource matching, temporal trends in female quality, or seasonal temperature increases (Perrins 1970, Verhulst and Nilsson 2008, Socolar et al. 2017). Regional temperature extremes have also been correlated both positively and negatively to daily nest survival in cavity-nesting species (Newlon and Saab 2011, Saab et al. 2011, Socolar et al. 2017). In burned forests, daily nest survival rates may decrease with increasing years since fire due to increases in nest predator populations, decreases in available snags, or decreases in prey density within snags that persist (Rota et al. 2014). While land managers generally cannot control abiotic/temporal variables, understanding how these factors relate to nest success can be important in making management decisions.
Burned forests are often targeted for snag removal to serve a variety of objectives, including hazard tree removal, economic gain, replanting, and reducing the risk of subsequent fire. These management actions have the potential to reduce populations of fire-associated species like the Black-backed Woodpecker (Hanson and North 2008, Tarbill et al. 2018), leading to conflict between management objectives after fire. Information on the factors that influence nest site selection and nest success, as well as descriptive information on the timing of breeding activities, can help managers reduce conflicts between wildlife-related objectives and other management needs.
In this study, we examined the effect of variation in post-fire habitat characteristics on Black-backed Woodpecker nesting ecology in California. Specifically, our objectives were to (1) identify important habitat components in burned forests that increase the probability of Black-backed Woodpecker nest site selection, and (2) evaluate the influence of habitat characteristics, nest characteristics, and abiotic/temporal factors (i.e. temperature, nest initiation date, day of nesting period) on Black-backed Woodpecker nest success. We predicted that the biotic factors with a strong influence on nest site selection would show adaptive significance by also affecting daily nest survival, as expected under the theory of adaptive habitat selection (Hildén 1965, Martin 1998). Alternatively, we evaluated the prediction that there would be a lack of congruence between the factors that influence nest site selection and nest success in post-fire forests. We sought to provide information to help guide conservation of Black-backed Woodpeckers in the context of post-fire forest management.
METHODS
Study Sites
We studied Black-backed Woodpecker nest sites within the perimeters of 6 wildfires that burned in Plumas and Lassen National Forests, California (Figure 1). The Peterson (burned in 2008) and Bald (2014) fires burned predominantly eastside pine forest comprised of Pinus ponderosa, P. jeffreyi, Calocedrus decurrens, and Juniperus occidentalis. The Moonlight (burned in 2007), Wheeler (2007), Sugarloaf (2009), and Chips (2012) fires burned primarily Sierran mixed-conifer forest dominated by P. ponderosa, Abies concolor, A. magnifica, Pseudotsuga menziesii, and C. decurrens (Supplemental Material Table S1). Study areas ranged from ~1,100 m to 2,100 m elevation and spanned 2–10 yr post-fire at the time of data collection. We collected data at some fires in up to 4 consecutive years (Supplemental Material Table S2).
Locations of the 6 wildfires where Black-backed Woodpecker nests were monitored in northern California.
Nest Searching and Monitoring
We searched for woodpecker nests from mid-April to late July, 2011–2018, using broadcast surveys to detect territorial individuals. Rather than systematically searching a small portion of each fire, we attempted to saturate as large an area as logistically feasible through exploratory searching, although we were unable to cover the full extent of each fire due to their large sizes (up to 30,897 ha) and limited road access. Surveyors broadcast Black-backed Woodpecker vocalizations and territorial drumming obtained from the Macaulay Library of Natural Sounds (Cornell Laboratory of Ornithology, Ithaca, New York, USA; G. A. Keller ML50117, ML50121, ML50124). Once detected, we attempted to follow individuals back to their nest site and used behavioral cues to locate the nest cavity. We also captured and radio-tagged a subset of adult Black-backed Woodpeckers (n = 49) as part of a separate study of home range size and habitat selection (Tingley et al. 2014, Stillman et al. 2019b), and we supplemented our nest searching by following tagged individuals to their nests. Radio-tagged individuals did not perceptibly alter their behavior around nests, and we found no evidence that radio-tags influenced adult or nest survival (apparent nest success rate was 0.82 and 0.71 for tagged and untagged birds, respectively). Additional information on capture and tagging methods can be found in Tingley et al. (2014).
We visited nests every 2–5 days (mean interval = 3.5 days) to record nest status and activity until fledging or failure. We assigned nest stage at each visit (laying, incubating, nestling) using adult and nestling behavioral cues during 15–60 min observation periods. We recorded nests as successful if they appeared to be empty on or near the expected fledging day and if nestling behavior during the previous visit suggested that nestlings would fledge soon (e.g., mature vocalizations, poking head out of cavity, advanced feather development). In many cases (59 out of 118 nests), we were able to confirm nest success with observations of recent fledglings in the vicinity. We recorded nest failure if the nest appeared empty or occupied by a different species well before the expected fledge date, or if there were clear signs of depredation (e.g., eggs crushed inside cavity, gnaw marks at cavity entrance). If we were unable to confidently assign nest fate (10 out of 118 nests), we excluded the last observation interval from our nest survival analysis.
Habitat Predictors of Nest Site Selection and Survival
We examined multiple nest, habitat, and abiotic/temporal variables (Table 1). Tree diameter and nest cavity orientation may regulate cavity temperature profiles with potential effects on survival (Hooge et al. 1999, Wiebe 2001), and nests that are higher may provide greater protection from ground-based predators (Fisher and Wiebe 2006). At a broader scale, we predicted that woodpeckers would place nests in pyrodiverse areas near the edges of high-severity patches in order to maintain proximity to preferred high-cover habitat for young fledglings while maintaining access to snag-dense foraging sites (Stillman et al. 2019b). Alternatively, we also evaluated whether nests near habitat edges (i.e. between high- and low-severity patches) would experience higher predation rates (Fisher and Wiebe 2006, Saab et al. 2011).
Covariates used in models of Black-backed Woodpecker daily nest survival. We grouped covariates into 3 categories while building the model set (nest characteristics, habitat characteristics, and abiotic/temporal factors) and explored different combinations of these covariate sets based on a priori hypotheses. We included nest characteristics and habitat characteristics in our analysis of nest site selection.
| Covariate set | Covariate name | Description |
|---|---|---|
| Nest characteristics | Tree DBH | Tree diameter at 1.4 m (cm) |
| Nest characteristics | Orientation | Orientation of the cavity entrance (°) |
| Nest characteristics | Cavity height | Height from ground to the cavity entrance (m) |
| Habitat characteristics | Distance to edge | Distance to the nearest low severity or unburned patch (m) |
| Habitat characteristics | Burn severity | The mean % change in canopy cover after fire, within 100 m |
| Habitat characteristics | Snag count | Number of snags >10 cm DBH within 10 m |
| Habitat characteristics | Time since fire | Number of years since the fire burned |
| Abiotic/temporal | Temp anomaly | Deviation from expected max temperature over the nesting period (°) |
| Abiotic/temporal | Initiation date | Julian day that the first egg was laid |
| Abiotic/temporal | Nest day | Day of the nesting period, ranging from 1 to 40 |
| Covariate set | Covariate name | Description |
|---|---|---|
| Nest characteristics | Tree DBH | Tree diameter at 1.4 m (cm) |
| Nest characteristics | Orientation | Orientation of the cavity entrance (°) |
| Nest characteristics | Cavity height | Height from ground to the cavity entrance (m) |
| Habitat characteristics | Distance to edge | Distance to the nearest low severity or unburned patch (m) |
| Habitat characteristics | Burn severity | The mean % change in canopy cover after fire, within 100 m |
| Habitat characteristics | Snag count | Number of snags >10 cm DBH within 10 m |
| Habitat characteristics | Time since fire | Number of years since the fire burned |
| Abiotic/temporal | Temp anomaly | Deviation from expected max temperature over the nesting period (°) |
| Abiotic/temporal | Initiation date | Julian day that the first egg was laid |
| Abiotic/temporal | Nest day | Day of the nesting period, ranging from 1 to 40 |
Covariates used in models of Black-backed Woodpecker daily nest survival. We grouped covariates into 3 categories while building the model set (nest characteristics, habitat characteristics, and abiotic/temporal factors) and explored different combinations of these covariate sets based on a priori hypotheses. We included nest characteristics and habitat characteristics in our analysis of nest site selection.
| Covariate set | Covariate name | Description |
|---|---|---|
| Nest characteristics | Tree DBH | Tree diameter at 1.4 m (cm) |
| Nest characteristics | Orientation | Orientation of the cavity entrance (°) |
| Nest characteristics | Cavity height | Height from ground to the cavity entrance (m) |
| Habitat characteristics | Distance to edge | Distance to the nearest low severity or unburned patch (m) |
| Habitat characteristics | Burn severity | The mean % change in canopy cover after fire, within 100 m |
| Habitat characteristics | Snag count | Number of snags >10 cm DBH within 10 m |
| Habitat characteristics | Time since fire | Number of years since the fire burned |
| Abiotic/temporal | Temp anomaly | Deviation from expected max temperature over the nesting period (°) |
| Abiotic/temporal | Initiation date | Julian day that the first egg was laid |
| Abiotic/temporal | Nest day | Day of the nesting period, ranging from 1 to 40 |
| Covariate set | Covariate name | Description |
|---|---|---|
| Nest characteristics | Tree DBH | Tree diameter at 1.4 m (cm) |
| Nest characteristics | Orientation | Orientation of the cavity entrance (°) |
| Nest characteristics | Cavity height | Height from ground to the cavity entrance (m) |
| Habitat characteristics | Distance to edge | Distance to the nearest low severity or unburned patch (m) |
| Habitat characteristics | Burn severity | The mean % change in canopy cover after fire, within 100 m |
| Habitat characteristics | Snag count | Number of snags >10 cm DBH within 10 m |
| Habitat characteristics | Time since fire | Number of years since the fire burned |
| Abiotic/temporal | Temp anomaly | Deviation from expected max temperature over the nesting period (°) |
| Abiotic/temporal | Initiation date | Julian day that the first egg was laid |
| Abiotic/temporal | Nest day | Day of the nesting period, ranging from 1 to 40 |
After nests fledged young or failed, we measured characteristics of nest trees and the habitat around each nest (Table 1). At the nest tree, we recorded 4 variables: diameter at breast height (DBH), cavity orientation, cavity height, and tree height. For habitat characteristics surrounding the nest, we counted the number of snags with DBH > 10 cm within a 10 m radius and collected 2 additional variables using remote sensing data. First, we used 30 m resolution Landsat-derived GIS data from the Rapid Assessment of Vegetation Condition after Wildfire program (https://www.fs.fed.us/postfirevegcondition/index.shtml) to extract the mean burn severity within 100 m of each nest tree, measured as the percent change in canopy cover from pre-fire to immediately after fire. Second, we measured the distance between the nest tree and the nearest forest patch burned at low severity or left unburned. We defined the threshold for “low severity” at ≤4.4% change in canopy cover, which is the median raster value of points classified as low severity from a field survey of ~10,000 points within the study area (Stillman et al. 2018). Nests located within a stand that burned at low severity (n = 4) were assigned a distance of zero.
Abiotic/Temporal Predictors of Daily Nest Survival
In addition to characteristics of nests and surrounding habitat, we also measured climatic and temporal variables with hypothesized ties to daily nest survival rate (Table 1). Given that our study sites were composed of open, snag-dominated habitats near the southern end of the species’ range, we predicted that high temperature anomalies at the regional scale would have a negative effect on woodpecker nest success (Saab et al. 2011, Socolar et al. 2017). We also predicted that nest success would be lower in late-season nesting attempts and that predation would be more likely when nests contained older, more vocal young (Verhulst and Nilsson 2008, Kozma and Kroll 2012).
We extracted daily maximum temperatures during the full 40-day period for each nest, regardless of nest fate, from the closest National Oceanic and Atmospheric Administration (NOAA) station to each nest (mean distance from nest to monitor = 10.6 km; data available at https://www.ncdc.noaa.gov). To calculate the daily anomaly in maximum temperature, we subtracted the mean maximum temperature experienced over the 40-day period from each daily value (Socolar et al. 2017).
We assigned an initiation date to each nest by counting backwards from observed transitions in nesting stage (laying, incubation, nestlings, fledglings) using published estimates of nesting period length (Tremblay et al. 2016). We assumed a 40-day full nesting cycle with 2 days for laying, 13 days for incubation, and 25 days between hatching and fledging, which proved consistent with our field observations. For successful nests, we estimated fledge date as the median date between the last nestling observation and the first post-fledging observation, and we back-counted 40 days to assign the initiation date. For failed or uncertain nest fates, we back-counted from our most confident nest transition date based on behavioral observations recorded during nest visits. Once each nest was given an initiation date, we assigned each interval between visits an ordinal day in the nesting period, from 1 to 40, by taking the median day between consecutive visits.
Sample of Available Nesting Locations
We evaluated nest site selection by comparing used nest locations to a sample of available nest locations at 2 spatial scales that are biologically relevant in our study system (Jones 2001). First, we evaluated nest stand selection at the home range scale by randomly selecting 5 “available” nest locations from an 806 m radius buffer around each nest. This buffer encompassed an area of 204 ha, which is the average minimum convex polygon home range size for Black-backed Woodpeckers in our study area (Tingley et al. 2014). This method allowed us to investigate third-order habitat selection—the selection of nest characteristics from within the individual’s home range (Johnson 1980). For each available location, we extracted the mean burn severity and the distance between the available point and the nearest low-severity or unburned edge, as above.
Second, we used field-based measurements to compare the used nest tree and available trees in a manner consistent with fourth-order habitat selection—the selection of specific resources from a resource patch (Johnson 1980). As part of 2 separate studies (Tingley et al. 2014, Stillman et al. 2019b), we established systematic 200 × 200 m grids of sampling points covering the minimum convex polygon of tracking locations for individual radio-tagged birds in each of the 6 fires. We used the subset of nests that overlapped the boundaries of sampling grids (n = 53) in our analysis of nest tree selection, and we generated an available sample by selecting the 5 closest grid points to each nest in the subset (mean distance ± SD = 137 ± 88 m). At each grid point, we measured the DBH of the closest snag and counted the number of snags with DBH > 10 cm within a 10 m radius of the point. We selected only snags (rather than live trees) for the available sample because the vast majority (98%) of our nests occurred in snags.
Nest Site Selection Analysis
We examined the biotic factors that influence nest site selection in 3 steps representing 3 spatial scales: (1) nest stand selection, (2) nest tree selection, and (3) nest cavity placement.
Nest stand selection
We used the full sample of nests to estimate relationships between nest stand selection and 2 GIS-derived landscape-level variables: burn severity and distance to low-severity or unburned edge. We used a generalized linear mixed effects model to estimate the parameters of the exponential resource selection function in a Bayesian framework. Our model included a random fire-level intercept term to account for potential nonindependence of nests within each fire (Gillies et al. 2006).
Nest tree selection
We used the subset of 53 nests that overlapped the systematic sampling grid to estimate relationships between nest tree selection and 2 field-measured variables: nest tree DBH and snag density around nest trees. We used a Bayesian generalized linear mixed effects model with a random intercept term to account for multiple samples in each fire-by-year combination. We included a quadratic term for DBH based on expectations from our field observations and Seavy et al. (2012).
Both nest selection models (stand and tree) contained 5 available locations for every nest location. We used vague normal prior distributions (μ = 0, τ = 0.1) on the intercept parameter and fixed effects, and a uniform (0.1, 3) prior distribution for the random effect standard deviation. We included initial values for all parameters based on a preliminary run of 1,200 iterations.
Nest cavity placement
We separately examined selection for 2 components of nest cavity placement: cavity orientation and cavity height above the ground. We tested for nonrandom cavity orientation using angular statistics and a Rayleigh test (Batschelet 1981). Given a sample of directional orientations, this test measures the amount of dispersion around the mean vector and uses the relative index (r) to test the null hypothesis that the orientations are randomly distributed. An r value of 1.0 indicates that all points have the same orientation and a value of 0 indicates random orientation. We tested for selection in nest cavity height using a bootstrapping approach with paired t-tests to compare observed heights to a series of “available” nest heights. We generated the available distribution by iteratively sampling 1,000 potential heights for each tree ranging from ground level to the measured tree height (for broken-top snags) or to 3 m below the measured tree height (for trees with intact tops). We used this method to avoid including portions of the tree trunk that were too thin to hold a potential cavity. For each of the 1,000 samples, we used a paired t-test to calculate the average difference between observed and random nest placements and made inference on the 95% confidence interval of this test statistic.
Nest Survival Models
We modeled Black-backed Woodpecker daily nest survival (the probability that a given nest survives from dayx to dayx+1) using logistic exposure models fit within a Bayesian framework (Schmidt et al. 2010). These models support the inclusion of fixed and random effects in a hierarchical structure that accounts for the inherent multilevel design of nest survival studies (i.e. multiple observations per nest). In addition, our models allow for uneven sampling intervals and inclusion of interval-specific covariates.
The survival of nest i over observation interval j was assumed to follow a Bernoulli distribution yi,j= Bernoulli(θ i,j) where yi,j = 1 if nest i survived interval j, yi,j = 0 if the nest failed, and θ i,j is the probability of nest i surviving interval j. Here, θ represents the daily survival probability, s, raised to the length of the risk exposure period such that , where ti,j is the length of interval j for nest i. We modeled the logit-transformed daily nest survival probability as a function of nest-level and interval-level covariates. We included a nest-level identifier as a random intercept in each model to account for multiple observations of each nest over the breeding period.
Model selection
We developed a set of 14 nest survival models based on a priori hypotheses for our system and purposefully sought to test all variables included in our nest site selection analysis. We considered 10 potential covariates comprising 3 categories: nest characteristics, habitat characteristics, and abiotic/temporal variables (Table 1). Rather than evaluate every possible combination of covariates, we built models with covariate combinations meant to test specific hypotheses about our system based on previous research (Forristal 2009, Saab et al. 2011, Kozma and Kroll 2012; Table 2, Supplemental Material Table S3).
Model selection results for 14 candidate models of Black-backed Woodpecker daily nest survival. Bayesian model selection employed leave-one-out cross-validation to calculate the information criteria LOOIC, presented here on the deviance scale. Models are ranked by ascending LOOIC values, with ΔLOOIC given as the difference between each candidate model and the top-ranking model.
| Model covariates | LOOIC | ΔLOOIC |
|---|---|---|
| Initiation date + Nest day | 285.16 | 0.00 |
| Initiation date + Nest day + Temp anomaly | 288.11 | 2.95 |
| Intercept only | 288.69 | 3.53 |
| Initiation date + Snag count + Burn severity + Distance to edge + Time since fire | 293.32 | 8.16 |
| Initiation date + Tree DBH + Cavity height + Orientation | 294.20 | 9.04 |
| Burn Severity + Time since fire | 295.20 | 10.04 |
| Tree DBH + Cavity height + Orientation | 295.46 | 10.30 |
| Cavity height + Distance to edge | 296.98 | 11.82 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire | 300.14 | 14.98 |
| Initiation date + Nest day + Temp anomaly + Tree DBH + Cavity height + Orientation | 301.52 | 16.36 |
| Cavity height + Distance to edge + Time since fire + Nest day | 302.19 | 17.03 |
| Snag count + Burn severity + Distance to edge + Time since fire | 304.28 | 19.12 |
| Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 305.73 | 20.57 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 309.59 | 24.43 |
| Model covariates | LOOIC | ΔLOOIC |
|---|---|---|
| Initiation date + Nest day | 285.16 | 0.00 |
| Initiation date + Nest day + Temp anomaly | 288.11 | 2.95 |
| Intercept only | 288.69 | 3.53 |
| Initiation date + Snag count + Burn severity + Distance to edge + Time since fire | 293.32 | 8.16 |
| Initiation date + Tree DBH + Cavity height + Orientation | 294.20 | 9.04 |
| Burn Severity + Time since fire | 295.20 | 10.04 |
| Tree DBH + Cavity height + Orientation | 295.46 | 10.30 |
| Cavity height + Distance to edge | 296.98 | 11.82 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire | 300.14 | 14.98 |
| Initiation date + Nest day + Temp anomaly + Tree DBH + Cavity height + Orientation | 301.52 | 16.36 |
| Cavity height + Distance to edge + Time since fire + Nest day | 302.19 | 17.03 |
| Snag count + Burn severity + Distance to edge + Time since fire | 304.28 | 19.12 |
| Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 305.73 | 20.57 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 309.59 | 24.43 |
Model selection results for 14 candidate models of Black-backed Woodpecker daily nest survival. Bayesian model selection employed leave-one-out cross-validation to calculate the information criteria LOOIC, presented here on the deviance scale. Models are ranked by ascending LOOIC values, with ΔLOOIC given as the difference between each candidate model and the top-ranking model.
| Model covariates | LOOIC | ΔLOOIC |
|---|---|---|
| Initiation date + Nest day | 285.16 | 0.00 |
| Initiation date + Nest day + Temp anomaly | 288.11 | 2.95 |
| Intercept only | 288.69 | 3.53 |
| Initiation date + Snag count + Burn severity + Distance to edge + Time since fire | 293.32 | 8.16 |
| Initiation date + Tree DBH + Cavity height + Orientation | 294.20 | 9.04 |
| Burn Severity + Time since fire | 295.20 | 10.04 |
| Tree DBH + Cavity height + Orientation | 295.46 | 10.30 |
| Cavity height + Distance to edge | 296.98 | 11.82 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire | 300.14 | 14.98 |
| Initiation date + Nest day + Temp anomaly + Tree DBH + Cavity height + Orientation | 301.52 | 16.36 |
| Cavity height + Distance to edge + Time since fire + Nest day | 302.19 | 17.03 |
| Snag count + Burn severity + Distance to edge + Time since fire | 304.28 | 19.12 |
| Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 305.73 | 20.57 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 309.59 | 24.43 |
| Model covariates | LOOIC | ΔLOOIC |
|---|---|---|
| Initiation date + Nest day | 285.16 | 0.00 |
| Initiation date + Nest day + Temp anomaly | 288.11 | 2.95 |
| Intercept only | 288.69 | 3.53 |
| Initiation date + Snag count + Burn severity + Distance to edge + Time since fire | 293.32 | 8.16 |
| Initiation date + Tree DBH + Cavity height + Orientation | 294.20 | 9.04 |
| Burn Severity + Time since fire | 295.20 | 10.04 |
| Tree DBH + Cavity height + Orientation | 295.46 | 10.30 |
| Cavity height + Distance to edge | 296.98 | 11.82 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire | 300.14 | 14.98 |
| Initiation date + Nest day + Temp anomaly + Tree DBH + Cavity height + Orientation | 301.52 | 16.36 |
| Cavity height + Distance to edge + Time since fire + Nest day | 302.19 | 17.03 |
| Snag count + Burn severity + Distance to edge + Time since fire | 304.28 | 19.12 |
| Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 305.73 | 20.57 |
| Initiation date + Nest day + Temp anomaly + Snag count + Burn severity + Distance to edge + Time since fire + Tree DBH + Cavity height + Orientation | 309.59 | 24.43 |
We compared the 14 fitted Bayesian models of nest survival using leave-one-out cross-validation to estimate the pointwise out-of-sample prediction accuracy (Vehtari et al. 2017). We employed an efficient computation of cross-validation that uses Pareto-smoothed importance sampling, implemented through the R package loo (Vehtari et al. 2018). The top-performing model was then selected using the difference in leave-one-out information criteria (LOOIC); this value is presented on the deviance scale, similar to other information criterion approaches. To corroborate our model selection results, we also calculated the widely applicable information criterion (WAIC; Watanabe 2010), a generalization of the Akaike information criterion, and confirmed that both model selection approaches selected the same top model (see Supplemental Material Table S3 for WAIC-based results).
For all nest survival models, we included an informed prior on the global intercept based on a previous study in burned mixed-conifer forest, which estimated the daily survival rate for Black-backed Woodpecker nests at 0.994 ± 0.2006 (Forristal 2009), giving a logit-scale prior distribution as Normal(μ = 5.109978, τ = 0.086). We used vague normal prior distributions (μ = 0, τ = 0.1) on all fixed effects and a uniform prior (0.1, 3) on the random effect. Data exploration indicated linear relationships with DBH and nest day, so we included both variables as linear terms rather than as quadratic predictors. We included time since fire in our group of habitat characteristics due to its notable effects on habitat structure (Smucker et al. 2005).
Bayesian Inference
We fit all Bayesian models using JAGS and the package R2jags in R (Su and Yajima 2015, R Core Team 2018). Models ran 3 chains of 20,000 iterations thinned by 10 with a burn-in of 5,000, yielding a final posterior sample of 4,500. We visually inspected traceplots to assess chain convergence and confirmed that the Gelman-Rubin statistic was <1.1 for every parameter estimate (Gelman et al. 2004). We centered and standardized continuous variables prior to modeling, and we present inference on parameters using 95% Bayesian credible intervals. The predictor variable for cavity orientation was centered by calculating the absolute value circular distance from the mean angle, so that orientations that deviate further from the mean had larger values.
To evaluate fit for the nest selection models, we created a test statistic based on the observed and predicted ratio of used to available points and conducted posterior predictive tests with associated Bayesian p-values (Gelman et al. 1996). Similarly, we conducted a posterior predictive check for the top nest survival model by calculating the Bayesian p-value for a test statistic based on the observed/predicted ratio of interval-level nest failures and nest successes. We considered Bayesian p-values less than 0.1 or greater than 0.9 as evidence for lack of fit (Gelman et al. 1996).
RESULTS
We located 118 Black-backed Woodpecker nests within the boundaries of the 6 wildfires and monitored nests for a total of 956 observation intervals (mean = 8 visits to each nest).
Nest Site Selection
The average burn severity, measured as the percent change in canopy cover within 100 m of nests, was 82 ± 2% SE, and the average distance between nests and the closest low-severity or unburned edge was 255 ± 21 m. Woodpeckers nested at sites with a mean of 15 ± 1 snags within 10 m of the nest tree (95% quantile = 2–31 snags), and the average nest tree had a DBH of 37 ± 1 cm (95% quantile = 22–67 cm). All but 2 nests (98%) occurred in dead trees.
Resource selection models showed significant positive selection for habitat patches that burned at high severity (Table 3, Figure 2A). For example, a high-severity burn patch with 100% change in canopy cover (i.e. all trees killed) was over 5 times more likely to be used as a nest site than a patch with 50% canopy cover change. However, woodpeckers also selected sites that were relatively close to low-severity or unburned edges, showing greater probability of nesting near high–low severity ecotones than in the center of large high-severity patches (Table 3, Figure 2B).
Parameter estimates for models of woodpecker nest stand selection, nest tree selection, and daily nest survival. Nest selection models estimated the relative probability of resource use based on Bayesian generalized linear mixed-effects models. The nest survival model is the top Bayesian logistic exposure model from multi-model comparison (Table 2). Estimates show means and 95% Bayesian credible intervals.
| Covariate | Parameter estimate |
|---|---|
| Nest stand selection model | |
| Intercept | −1.83 (−2.66, −0.86) |
| Burn severity (% change in canopy cover) | 1.76 (1.34, 2.18) |
| Distance to edge | −0.96 (−1.40, −0.57) |
| Standard deviation of random fire effect | 0.94 (0.26, 2.29) |
| Nest tree selection model | |
| Intercept | −1.52 (−2.12, −0.97) |
| Snag count | 1.36 (0.95, 1.78) |
| Tree DBH | 2.20 (1.44, 3.08) |
| Tree DBH2 | −1.28 (−1.95, −0.75) |
| Standard deviation of random fire-year effect | 0.41 (0.11, 1.13) |
| Top nest survival model | |
| Intercept | 5.60 (4.70, 7.00) |
| Initiation date | −0.71 (−1.43, −0.17) |
| Nest day | −0.27 (−0.94, 0.26) |
| Standard deviation of random nest effect | 1.43 (0.21, 2.86) |
| Covariate | Parameter estimate |
|---|---|
| Nest stand selection model | |
| Intercept | −1.83 (−2.66, −0.86) |
| Burn severity (% change in canopy cover) | 1.76 (1.34, 2.18) |
| Distance to edge | −0.96 (−1.40, −0.57) |
| Standard deviation of random fire effect | 0.94 (0.26, 2.29) |
| Nest tree selection model | |
| Intercept | −1.52 (−2.12, −0.97) |
| Snag count | 1.36 (0.95, 1.78) |
| Tree DBH | 2.20 (1.44, 3.08) |
| Tree DBH2 | −1.28 (−1.95, −0.75) |
| Standard deviation of random fire-year effect | 0.41 (0.11, 1.13) |
| Top nest survival model | |
| Intercept | 5.60 (4.70, 7.00) |
| Initiation date | −0.71 (−1.43, −0.17) |
| Nest day | −0.27 (−0.94, 0.26) |
| Standard deviation of random nest effect | 1.43 (0.21, 2.86) |
Parameter estimates for models of woodpecker nest stand selection, nest tree selection, and daily nest survival. Nest selection models estimated the relative probability of resource use based on Bayesian generalized linear mixed-effects models. The nest survival model is the top Bayesian logistic exposure model from multi-model comparison (Table 2). Estimates show means and 95% Bayesian credible intervals.
| Covariate | Parameter estimate |
|---|---|
| Nest stand selection model | |
| Intercept | −1.83 (−2.66, −0.86) |
| Burn severity (% change in canopy cover) | 1.76 (1.34, 2.18) |
| Distance to edge | −0.96 (−1.40, −0.57) |
| Standard deviation of random fire effect | 0.94 (0.26, 2.29) |
| Nest tree selection model | |
| Intercept | −1.52 (−2.12, −0.97) |
| Snag count | 1.36 (0.95, 1.78) |
| Tree DBH | 2.20 (1.44, 3.08) |
| Tree DBH2 | −1.28 (−1.95, −0.75) |
| Standard deviation of random fire-year effect | 0.41 (0.11, 1.13) |
| Top nest survival model | |
| Intercept | 5.60 (4.70, 7.00) |
| Initiation date | −0.71 (−1.43, −0.17) |
| Nest day | −0.27 (−0.94, 0.26) |
| Standard deviation of random nest effect | 1.43 (0.21, 2.86) |
| Covariate | Parameter estimate |
|---|---|
| Nest stand selection model | |
| Intercept | −1.83 (−2.66, −0.86) |
| Burn severity (% change in canopy cover) | 1.76 (1.34, 2.18) |
| Distance to edge | −0.96 (−1.40, −0.57) |
| Standard deviation of random fire effect | 0.94 (0.26, 2.29) |
| Nest tree selection model | |
| Intercept | −1.52 (−2.12, −0.97) |
| Snag count | 1.36 (0.95, 1.78) |
| Tree DBH | 2.20 (1.44, 3.08) |
| Tree DBH2 | −1.28 (−1.95, −0.75) |
| Standard deviation of random fire-year effect | 0.41 (0.11, 1.13) |
| Top nest survival model | |
| Intercept | 5.60 (4.70, 7.00) |
| Initiation date | −0.71 (−1.43, −0.17) |
| Nest day | −0.27 (−0.94, 0.26) |
| Standard deviation of random nest effect | 1.43 (0.21, 2.86) |
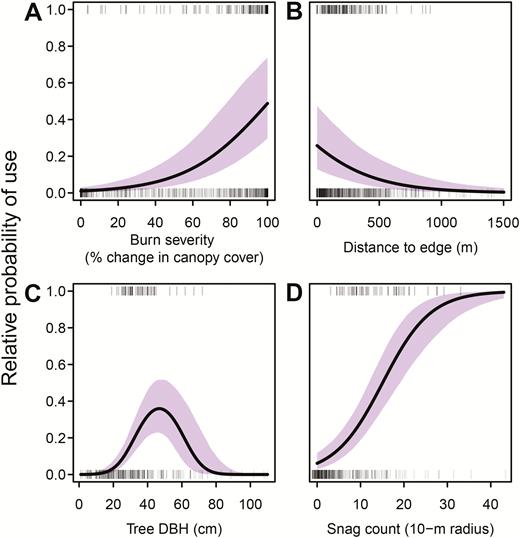
Modeled relationships for Black-backed Woodpecker nest stand selection (A, B) and nest tree selection (C, D). Thick black lines show the mean response predicted by the model, and shaded regions show 95% CI. Rugs depict the raw data, with used nest locations drawn at y = 1 and available locations drawn at y = 0.
We used data from a subset of 53 nests with field-based systematic habitat sampling to estimate the effect of tree diameter and snag density on nest tree selection. Woodpeckers showed a strong quadratic relationship to tree DBH, with peak probability of use at 47 cm (Table 3, Figure 2C). As snag density increased, the relative probability of nest tree use also increased (Figure 2D). Nest sites with 15 snags within a 10 m radius (equal to a density of ~5 snags per 100 m2) had a 50% relative probability of use.
Nest cavities showed a strong bias in orientation, with nonrandom clustering around the circular mean orientation of 80.6° (Rayleigh r = 0.36, P < 0.001; Figure 3A). The average nest cavity height was 4.8 ± 0.4 m (Figure 3B), which was significantly lower by 1.2 m (95% CI: −1.9, −0.5) than would be expected if nest height was uniformly random.
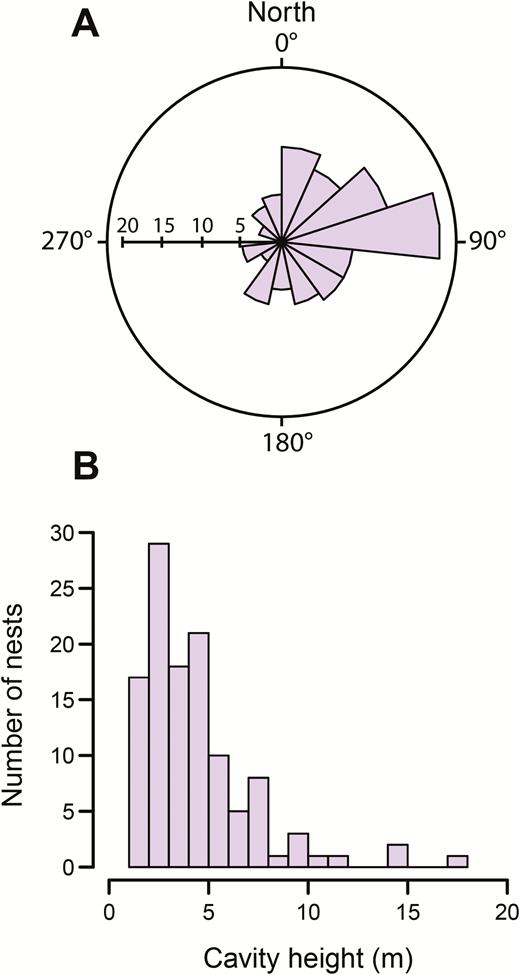
Nest cavity characteristics for 118 Black-backed Woodpecker nests in northern California, showing (A) the direction that nest cavities faced with sample size on the horizontal axis, and (B) a histogram of nest cavity heights.
Nest Survival
We observed nest failure at 27 of 108 nests with known fates, yielding an apparent nest success rate of 0.75. Though we were not able to confidently determine the cause of failure for most nests, we documented probable occurrences of nest predation by small mammals, 2 occasions of nest predation by American black bears (Ursus americanus), and one occasion of nest eviction by European Starlings (Sturnus vulgaris). We recorded failures for 3 nests during the laying stage, 12 during incubation, and 12 during the nestling stage. The median nest initiation date was May 3, and 75% of nests were initiated between April 21 and May 20 (Figure 4A). For documented successful nests (n = 81), the median fledge date was June 8 and 75% of successful nests fledged between May 30 and June 23 (Supplemental Material Figure S1).
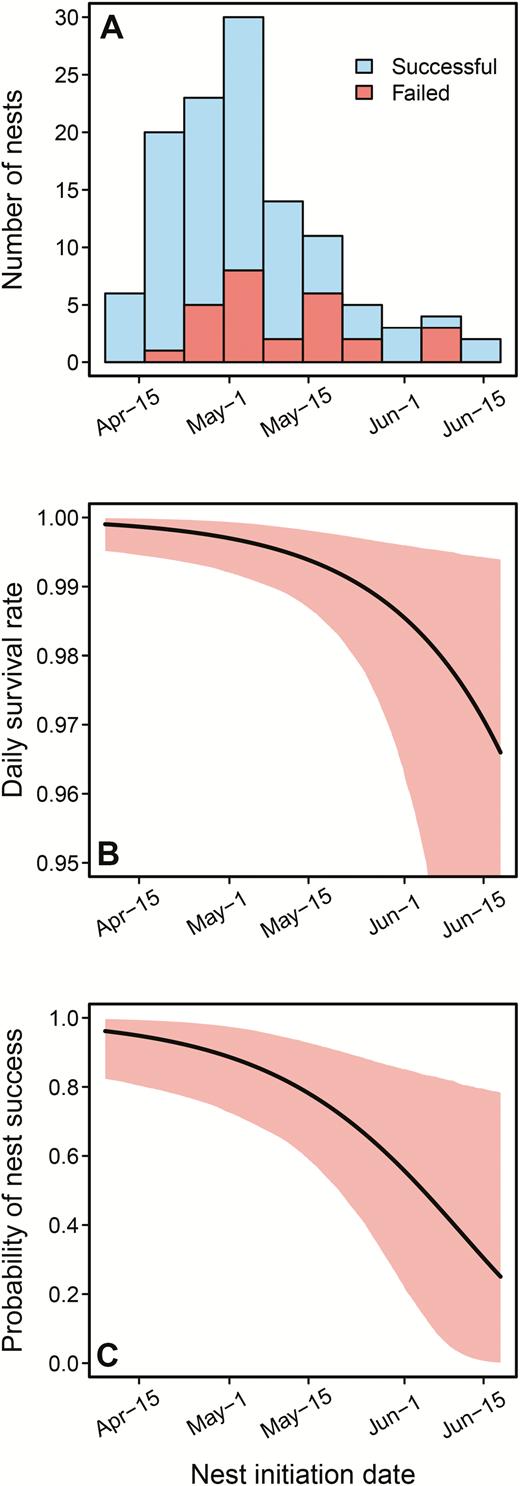
(A) Summary of nest initiation dates from 118 Black-backed Woodpecker nests in post-fire forests, split between successful and failed nests. Line plots show (B) predicted daily survival rate and (C) overall probability of nest success as a function of nest initiation date. Mean relationships are shown as black lines and shaded regions depict 95% CI. Predictions are shown from the best-supported nest survival model.
We evaluated 14 Bayesian nest survival models with 10 potential covariates. The top model for nest survival included initiation date and nest day (Table 2), but only initiation date showed a strong effect on nest survival (Table 3, Figure 4). Models that fit better than the intercept-only model included abiotic variables alone (Table 2), without any factors related to habitat or nest characteristics. Model estimates indicated that woodpecker nests that began later in the breeding season had a lower probability of survival, with daily survival rate dropping precipitously for nests initiated after May 20 (Figure 4B). Under average conditions, the top model estimated baseline daily survival rate at 0.996 (95% CI: 0.991, 0.999) and overall nest success rate at 0.852 (95% CI: 0.697, 0.961).
Model Fit
Both nest selection models closely approximated a test statistic derived from the expected number of used versus available locations (Supplemental Material Figure S2). Bayesian p-values indicated no significant lack of fit, with p = 0.49 for the model of nest stand characteristics and p = 0.48 for the model of tree characteristics. Similarly, our posterior predictive test on the expected ratio of nest successes vs. failures found no evidence for lack of fit in the top nest survival model (Bayesian p = 0.48).
DISCUSSION
In our study, Black-backed Woodpeckers showed strong habitat selection at the scale of nest stand, nest tree, and nest placement by narrowly choosing nesting locations relative to available habitat. However, variation in these selected habitat conditions across monitored nests showed no evidence for an effect on nest survival; rather, abiotic/temporal variables alone, and nest initiation date in particular, provided the best predictors of nest survival. Consequently, Black-backed Woodpecker nesting ecology in burned forests of northern California demonstrated a pattern of neutral congruence between the habitat characteristics important to nest site selection and nest survival.
Although theory predicts that nest site selection should show adaptive significance, our observational study did not detect a significant effect of habitat on nest survival. It is plausible that the main selective pressures operating in our study system (i.e. predation risk) were random or unpredictable within the range of selected habitats. This explanation is consistent with the adaptive peaks hypothesis (Wright 1932, Latif et al. 2012), which suggests that some bird populations may only nest in habitat that is already “adaptive,” and thus may not express sufficient variation in habitat selection necessary to detect links between fitness and habitat. Alternatively, nest site selection may respond primarily to other fitness components that were not part of our study, such as adult condition, adult survival, clutch size, or fledgling survival. Some aspects of nest site selection may also respond primarily to availability rather than apparent fitness benefits. For example, Black-backed Woodpeckers may select specific nest cavity locations based solely on the availability of suitable excavation sites on the nest tree instead of perceived predation risk (Lorenz et al. 2015).
Black-backed Woodpeckers are known to occur in low densities in unburned forests throughout the species’ range (Fogg et al. 2014, Tremblay et al. 2015a), but our study focused entirely on birds detected inside the perimeters of recent fires. Previous research elsewhere in the species’ range suggests that reproductive success is lower outside of wildfire perimeters (Rota et al. 2014), and populations within recently burned forest may act as source populations for the surrounding region (Nappi and Drapeau 2009). Future work directly comparing reproductive success in burned vs. unburned habitats would be beneficial (Tremblay et al. 2015a).
Post-fire logging has been demonstrated to reduce Black-backed Woodpecker abundance, likely by reducing the amount of available habitat for nesting and foraging (Saab et al. 2007, Hanson and North 2008). Post-fire logging treatments at the fires that we sampled included a broad spectrum of actions, ranging from limited roadside hazard tree removal to stand harvest for biofuel. While we attempted to prioritize nest searching in intact stands, our sample included some nests near previously logged areas. One nest was located in an area of active logging; management personnel retained a small cluster of trees around the nest, which fledged successfully, although both fledglings from this nest were depredated soon after fledging (A. N. Stillman, personal observation). While we did not test effects of post-fire logging on nest site selection or survival, the presence of these operations could have decreased the density of breeding Black-backed Woodpeckers in some study areas and may have caused some birds to select habitat outside of affected stands.
Nest Site Selection
Woodpeckers tended to select nest locations in high burn severity patches (Figure 2A), but typically within 500 m of low-severity patches or unburned edge (Figure 2B). Recently fledged Black-backed Woodpeckers generally move out of high-severity burn stands to seek greater cover (Stillman et al. 2019b), and we speculate that selection for nest sites closer to edges of high-severity patches provides easier access to preferred fledgling habitat, which may increase fledgling survival. Furthermore, proximity to fire edges or low-severity patches may have direct consequences on nest success, although we did not detect these effects in our study. In Quebec, Black-backed Woodpecker nests in burned forests showed increased productivity closer to unburned edges (Nappi and Drapeau 2009), perhaps due to increased concentrations of wood-boring beetle larvae (Monochamus scutellatus) in these areas (Saint-Germain et al. 2004). However, in Idaho, Saab et al. (2011) found that nest success decreased closer to unburned edges, perhaps because unburned forests act as a source for nest predators. Apparent differences in the effect of distance to edge on Black-backed Woodpecker nesting ecology may stem from regional differences in predator communities, forest types, or fire severity patterns. While we did not detect any relationship between nest survival and distance to edge at our study sites in northern California, we note that our woodpeckers nested farther from the edge on average than nests recorded by Saab et al. (2011) (mean distance to unburned or low-severity edge in California = 255 m, mean distance to unburned edge in Idaho = 159 m).
Although Black-backed Woodpeckers are known to preferentially forage on larger-diameter trees (Nappi et al. 2003), the birds in our study selected moderate-sized trees for nests, consistent with previous research in northern California (Seavy et al. 2012). Smaller-diameter trees (i.e. <15 cm DBH) are likely too thin to allow for cavity excavation, and larger-diameter trees may have comparatively reduced internal wood softening from wood-rotting fungi. When selecting excavation sites, Black-backed Woodpeckers may respond primarily to wood hardness rather than external tree characteristics (Lorenz et al. 2015). Given this, it is reasonable to expect regional discrepancies in favored nest tree characteristics due to geographic variation in fungal communities as well as the species and size of available trees. For example, Black-backed Woodpeckers in South Dakota selected relatively smaller nest trees with an average DBH of 25 cm (Bonnot et al. 2009), whereas birds in our northern California sites selected moderate-sized trees with an average DBH of 37 cm.
Nest cavities were lower than expected by chance, despite potential threats from ground-based predators. In addition to probable nest predation from small mammals, we documented 2 occurrences of nest predation by American black bears, which climbed trees to forcibly chew and claw a large opening at the cavity entrance. Although we did not detect an effect of nest height on survival, we hypothesize that higher nests may incur additional risks that balance nest placement toward the moderately low heights that we observed, such as an increased risk of snags snapping and falling at a point below the cavity.
Nest Survival in Selected Habitats
Despite the strong effect of habitat on nest selection, nest initiation date was the only significant predictor of daily nest survival probability. Reproductive success declines over the course of the breeding season for many bird species, although the direct mechanisms leading to this phenomenon are often complex (Verhulst and Nilsson 2008, Kozma and Kroll 2012). These seasonal patterns may stem either from variation in environmental quality (e.g., food availability, temperature extremes, predator abundance) or seasonal trends in the quality of breeding individuals (e.g., high-quality females breed first) (Perrins 1970). In our study system, we propose 2 potential mechanisms for the initiation–survival relationship. First, increased failure in later nests may result from breeding attempts by lower-quality individuals that renested after initial nest failures. Black-backed Woodpeckers generally raise 1 brood per breeding season, but they will excavate a second nest in a different tree if the first nest fails (Tremblay et al. 2016). We observed that 5 of 6 confirmed renesting attempts failed, and we suspect that some additional late nesting attempts in our sample came from renesting birds. Second, later nests may be exposed to increased risk of predation by nest predators that show temporal trends in nest predation rates (Fisher and Wiebe 2006).
Although past studies have demonstrated an effect of temperature on woodpecker nest success (Newlon and ,Saab 2011, Saab et al. 2011), temperature variation at the regional scale did not have a direct effect on daily nest survival in our study area. This result could arise from not directly measuring temperature at active nests, as cavity-nesting birds can potentially mitigate the effects of temperature extremes by selecting appropriate microhabitat characteristics. East-facing cavities, which represented the majority of our 118 nests, are thought to mediate temperature fluctuations in cavity-nesting species—nests that face east average higher morning temperatures and cooler afternoon temperatures than nests that face other directions (Hooge et al. 1999, Tremblay et al. 2015b).
Our results did not support our prediction that Black-backed Woodpecker nest success decreases with increasing time since fire. Although we sampled some fires in multiple consecutive years, we primarily employed a space-for-time substitution in order to build a sample of fires ranging from 2 to 10 yr post-fire. Because fires differed in elevation, latitude, and forest composition, it is possible that environmental differences between study sites masked a true decline in survival with increasing time since fire. However, the strong relationship between habitat and nest site selection suggests that, even >7 yr post-fire, woodpeckers may still maintain high-quality nest sites by selecting for remaining stands of dense snags in high-severity burn patches. Thus, the gradual decrease in snag density or prey abundance with time since fire may show a stronger effect on woodpecker abundance than on nest success. In this scenario, decreases in suitable habitat would cause a decrease in abundance even as nest success remains constant. Previous research has repeatedly demonstrated temporal declines in Black-backed Woodpecker occupancy, population persistence, and nest density with increasing time since fire (Saab et al. 2007, Tingley et al. 2018, Matseur et al. 2019), with conflicting evidence for coincidental declines in nest survival (Nappi and Drapeau 2009, Rota et al. 2014).
Management Implications
Fire regimes in the American West are increasingly characterized by large, homogeneously severe wildfires, prompting concern for the conservation of forest bird populations (Miller and Safford 2012, Tingley et al. 2016, Stevens et al. 2017). It may seem that post-fire specialists like the Black-backed Woodpecker stand to benefit from an increased prevalence of high-severity post-fire landscapes in California, but recent research suggests that this species seeks out more pyrodiverse areas—landscapes that have greater variation in burn severity—to meet the habitat preferences of both fledglings and adults (Stillman et al. 2019b). The finding that the woodpeckers in our study tended to select nest sites closer to the edges of high-severity burn patches provides added evidence that pyrodiversity is an important component of Black-backed Woodpecker habitat within burned forests, consistent with research in other parts of the species’ range (Nappi et al. 2010). We suggest that managers interested in providing nesting habitat for Black-backed Woodpeckers consider retention and creation of pyrodiverse landscapes that include dense stands of snags (>5 snags/100 m2) within ~500 m of stands that burned at low severity or remained unburned.
Managers may reduce conflicts between wildlife conservation and objectives that involve snag removal by shifting the timing of habitat-altering activities. Our finding that nest success declines in later-season nests suggests that managers could reduce the immediate impact of logging activities on breeding Black-backed Woodpeckers by delaying actions until after most nests have fledged. At our study sites in northern California, 95% of nests had fledged by June 29, which may provide a useful benchmark for timing post-fire management that might disturb nesting habitat.
More generally, our results provide a caution for land managers interested in animal conservation: when a species exhibits strong behavioral selection for high-quality habitat, purely observational studies may fail to reveal relationships between habitat and fitness (Chalfoun and Schmidt 2012). In this scenario, abiotic/temporal variables—which are generally not controlled by land managers—will likely have the strongest effect on fitness components like nest success. We might expect this phenomenon to be especially prevalent in habitat specialists, where natural selection has presumably shaped behavioral habitat selection to reflect a narrow range of preferred conditions. When suitable habitat is abundant, habitat specialists can behaviorally select nest locations that confer a relative fitness advantage, and thus avoid the negative consequences of breeding in poor-quality areas.
ACKNOWLEDGMENTS
We thank our dedicated field technicians for their reliable and persistent hard work, and we are grateful to M. Bond, N. Hagemeyer, K. Hein Strohm, M. Lerow, and F. Tousley for leadership in the field. We also thank D. Craig for facilitating support, and R. Burnett for assistance in locating Black-backed Woodpecker nests. Manuscript drafts benefitted substantially from guidance provided by C. Elphick, M. Reed, and T. Rittenhouse. This is Contribution No. [630] of The Institute for Bird Populations.
Funding statement: This work was supported by Plumas National Forest and the Pacific Southwest Region of the US Forest Service. Additional funding was provided by the American Ornithological Society Hesse Award, the American Philosophical Society Lewis and Clark Fund for Exploration and Field Research, the Wilson Ornithological Society Paul A. Stewart Grant, the Sierra Foothills Audubon Society, and the UConn EEB Zoology Award (2016–2018).
Ethics statement: This research was conducted in compliance with the University of Connecticut Institutional Animal Care and Use Committee (IACUC protocol #A16-008), the California Department of Fish and Wildlife (Scientific Collecting Permit #SC-8645), and the USFWS Bird Banding Laboratory (Permit #22423).
Author contributions: RBS, RLW, MWT, and ANS conceived the ideas and designed methodology; RLW, ANS, and RBS oversaw data collection with support from MJ and CAH; ANS and MWT analyzed the data; and ANS led writing of the first draft of the manuscript. All authors contributed to manuscript drafts and gave final approval for publication.
Data depository: Analyses reported in this article can be reproduced using the data provided by Stillman et al. (2019a). Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.n98q8s3

{kind=link}
{kind=link}
{kind=link}
{kind=link}