
Seagrass and Oyster Reef Restoration in Living Shorelines: Effects of Habitat Configuration on Invertebrate Community Assembly

Abstract
:1. Introduction
2. Materials and Methods
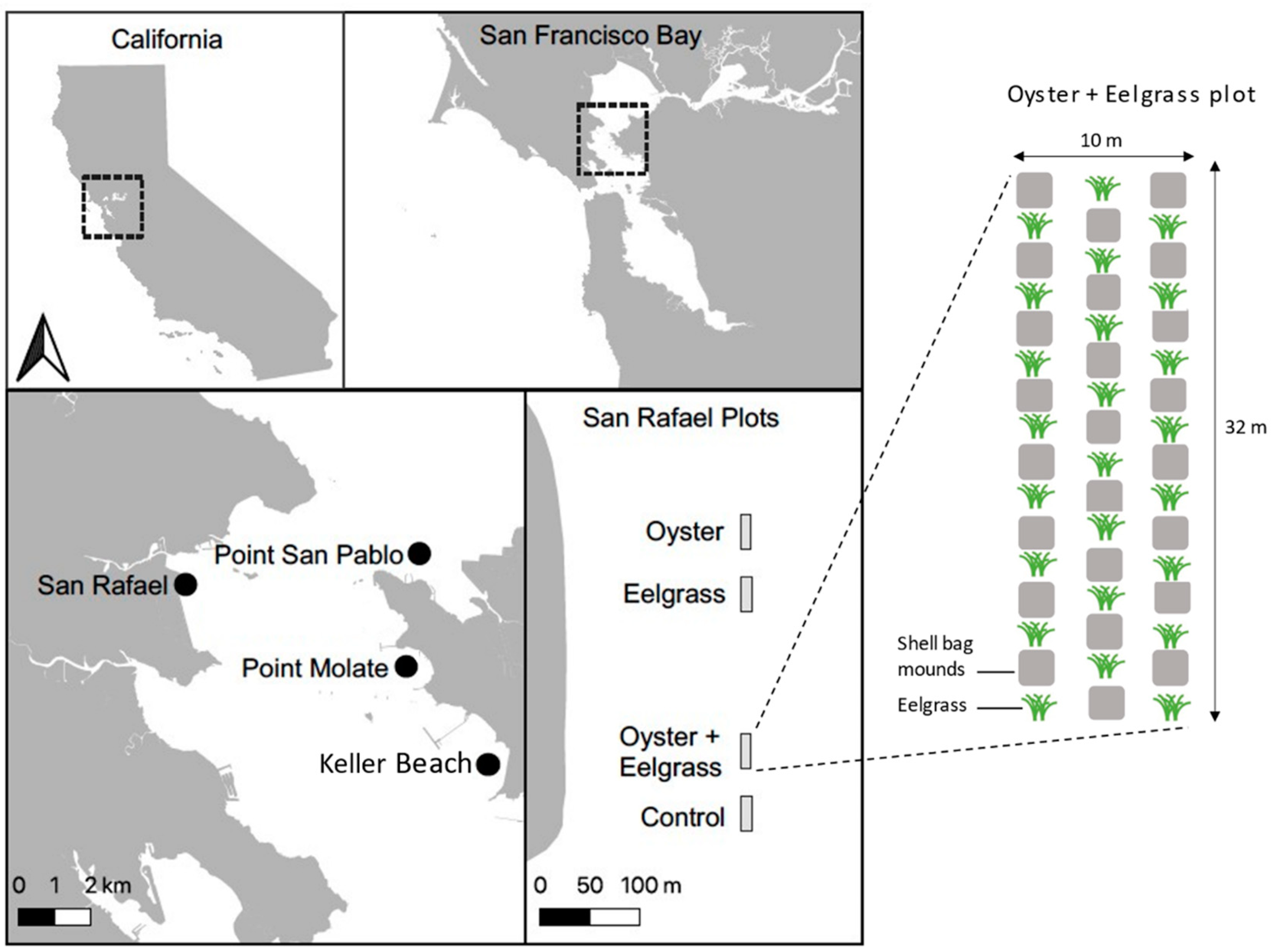
2.1. Location
2.2. Treatment Design
2.3. Monitoring
2.3.1. Suction Sampling
2.3.2. Eelgrass Shoot Sampling
2.4. Data Analysis
3. Results
3.1. Eelgrass and Oyster Reef Habitat Development
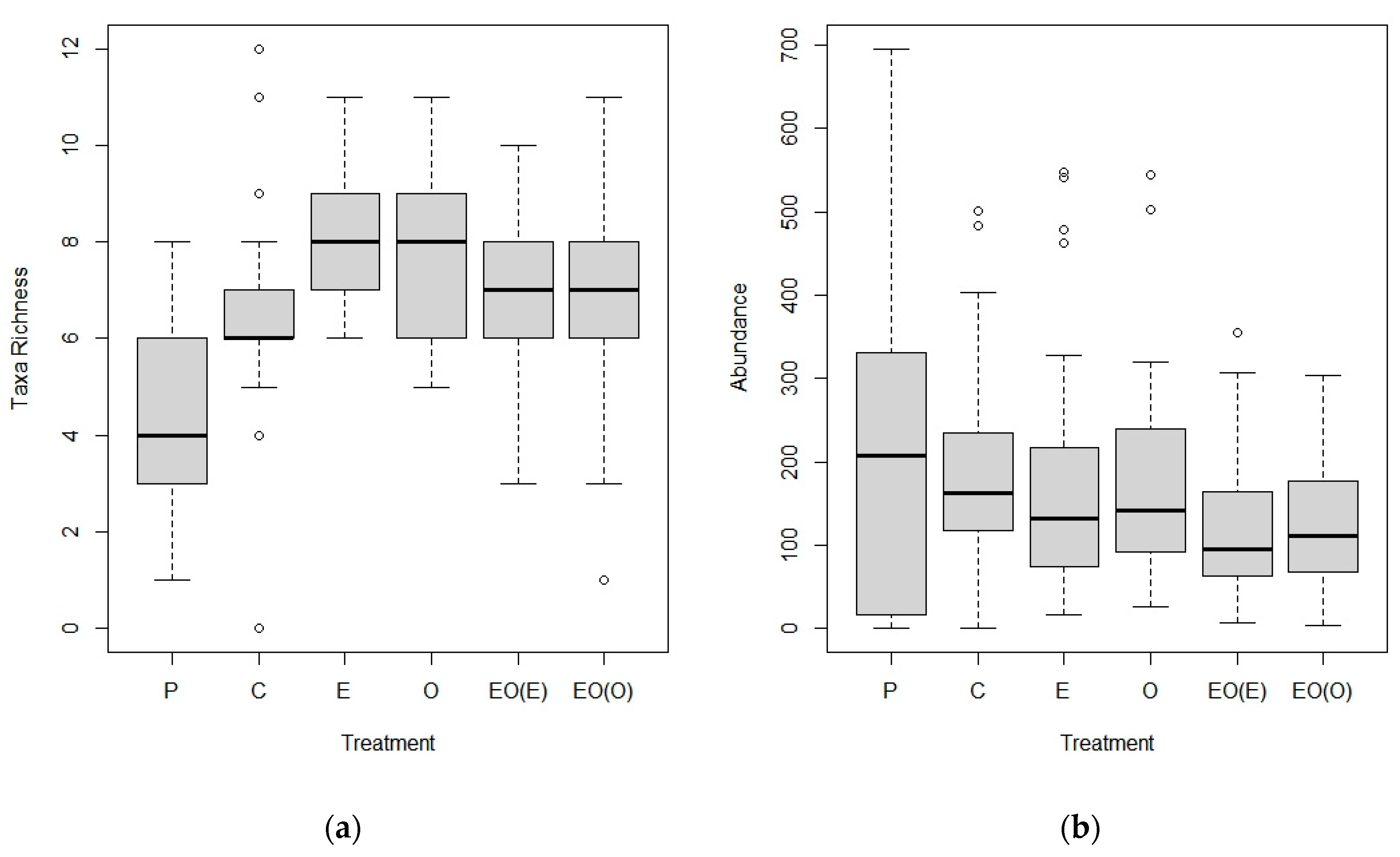
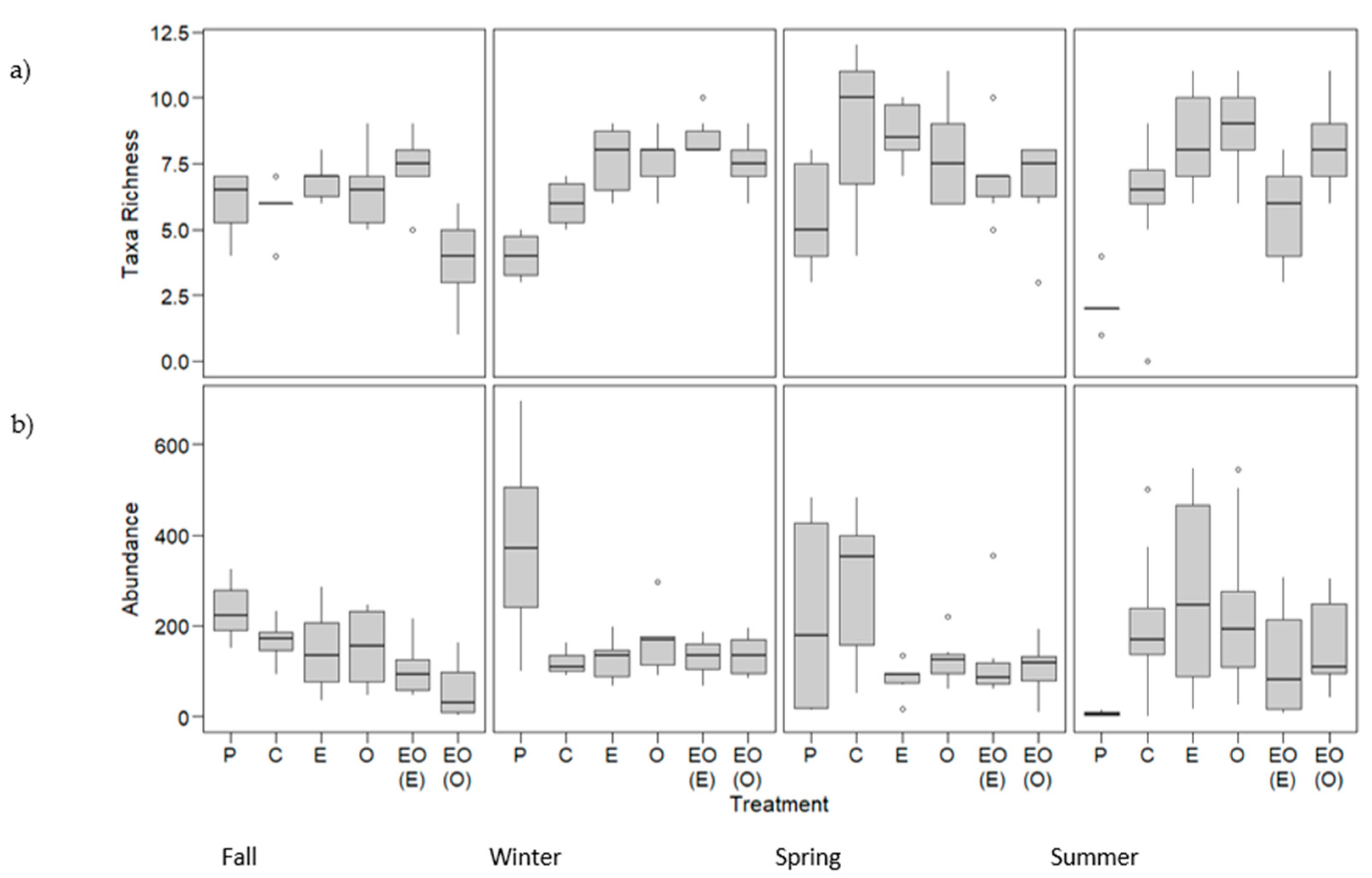
3.1.1. Taxa Richness and Abundance: Suction Sampling
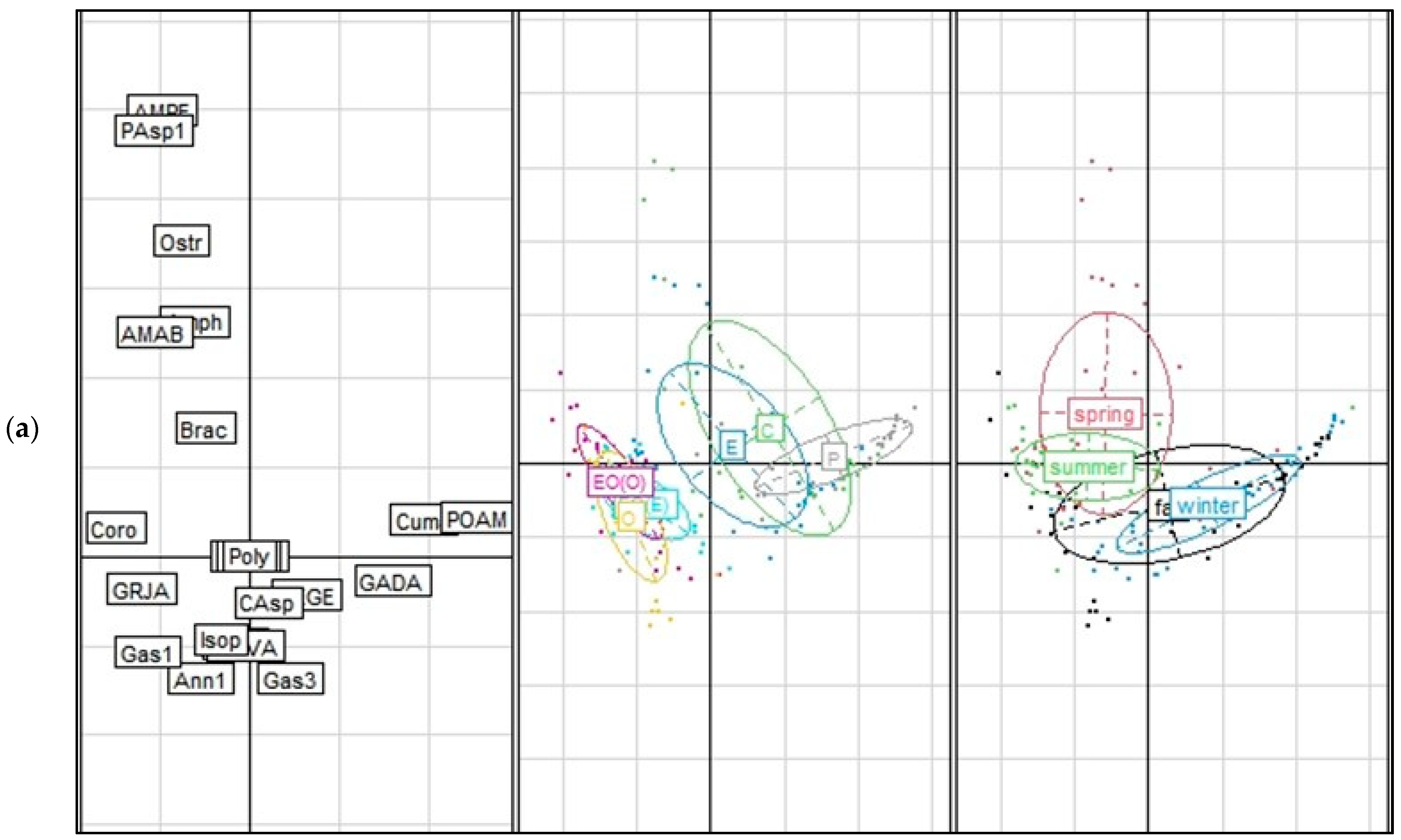
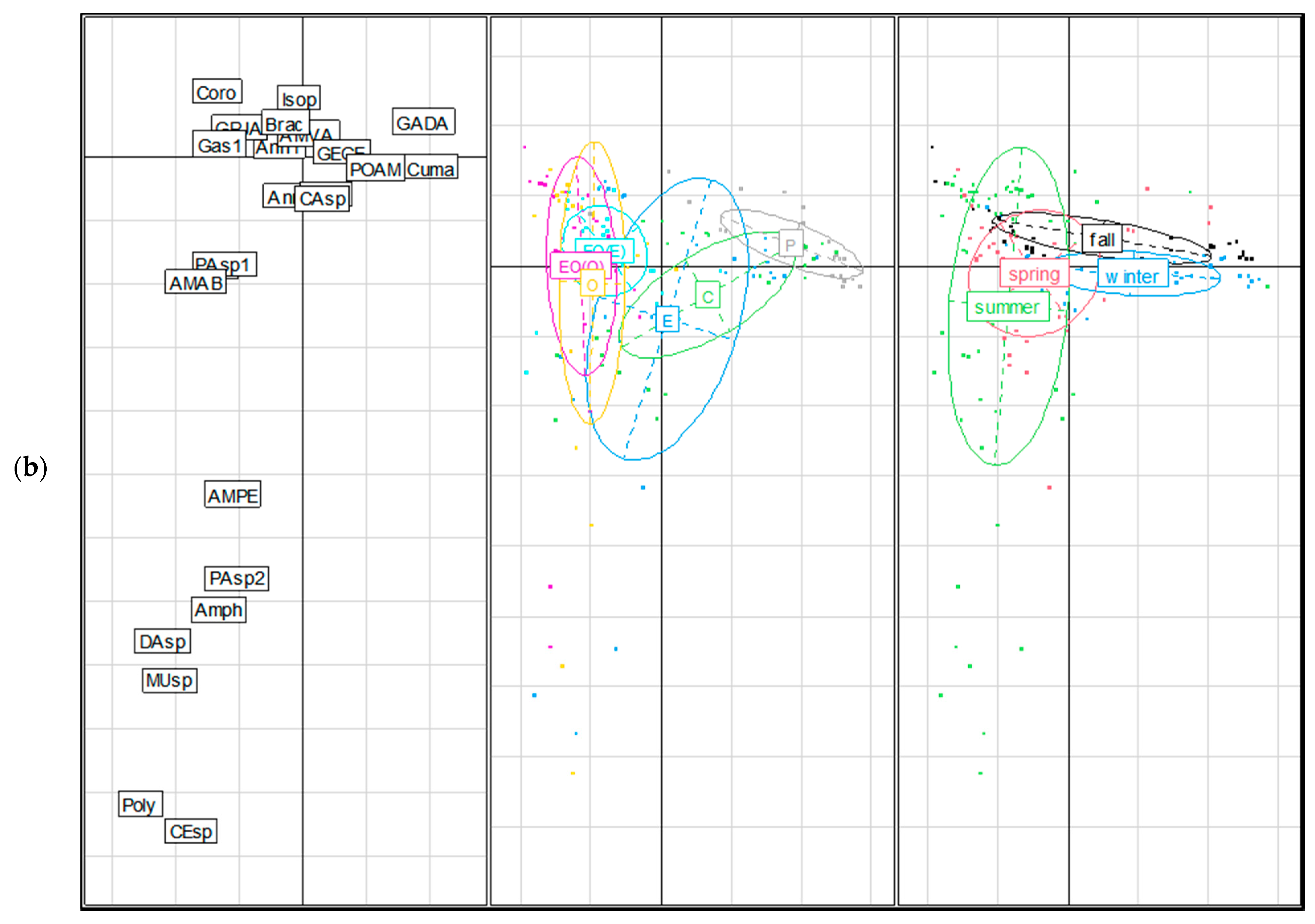
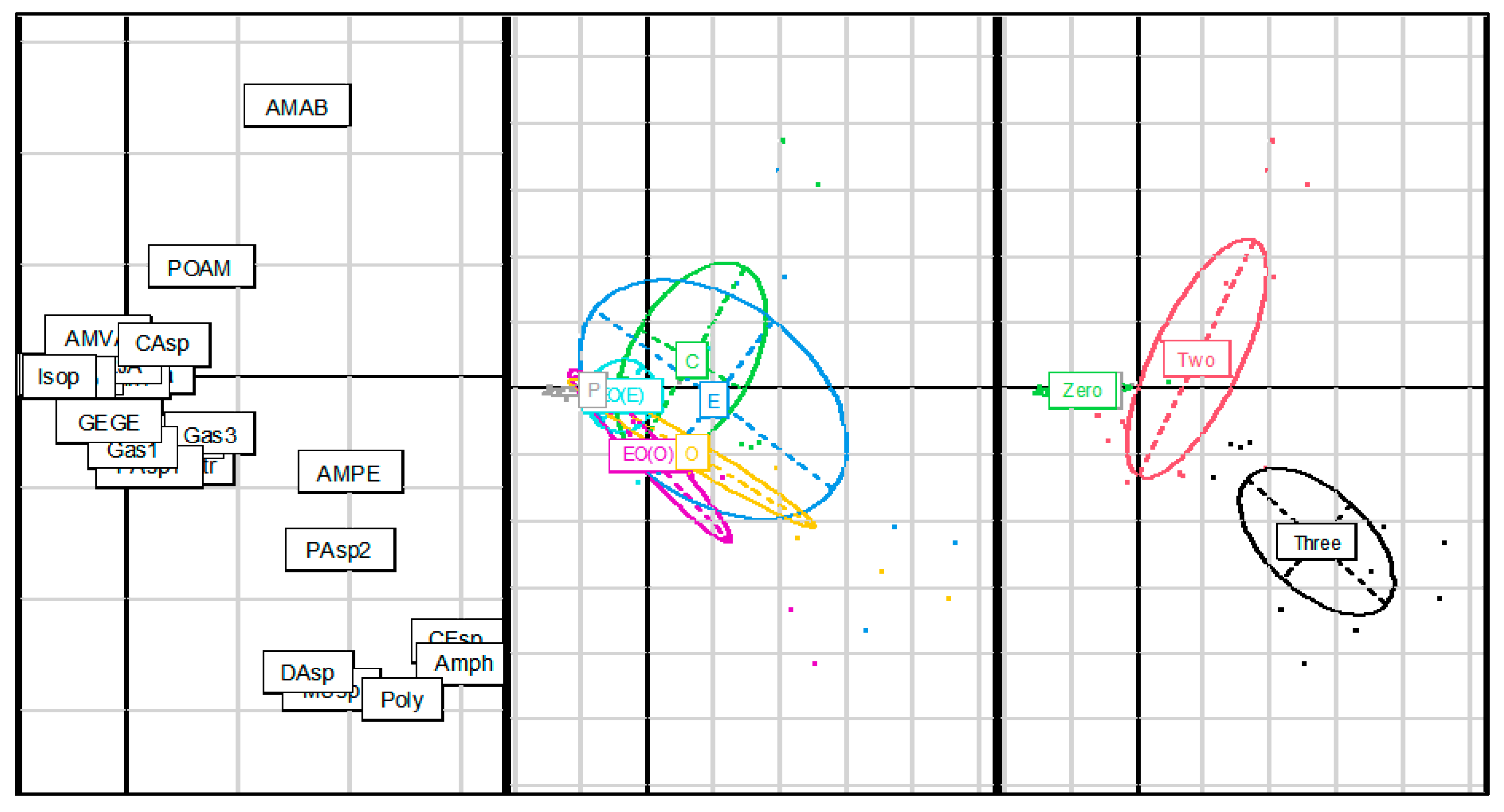
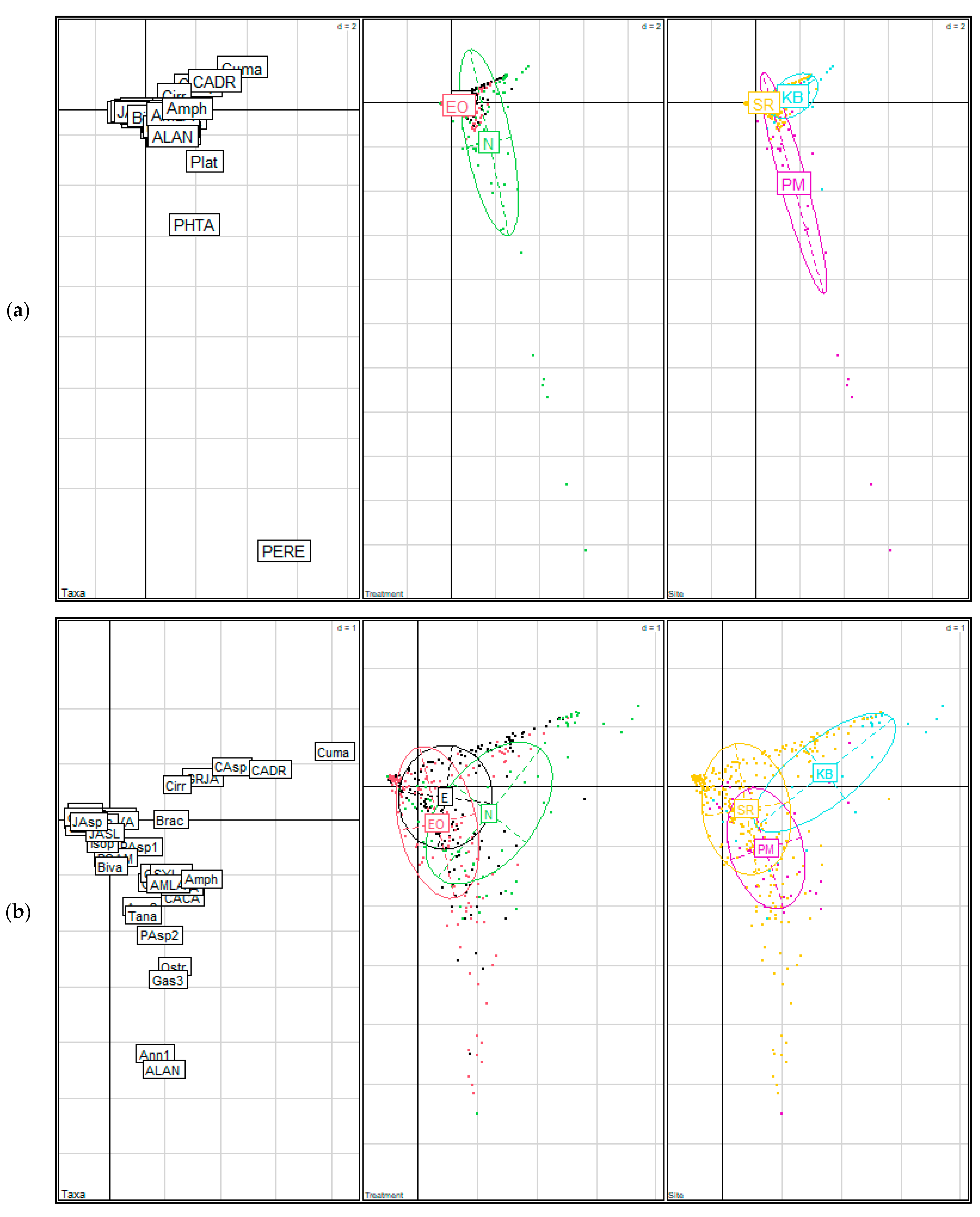
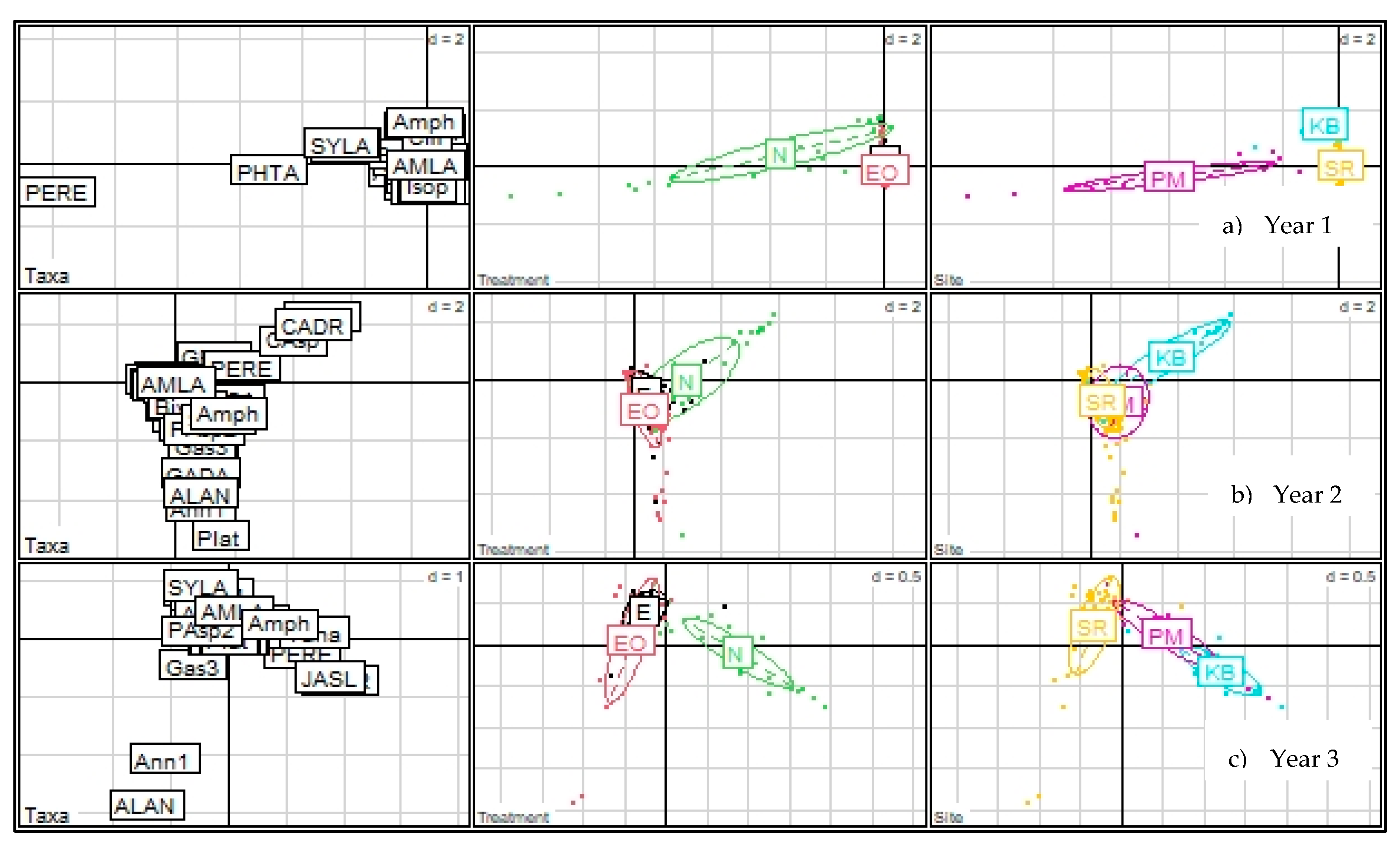
3.1.2. Community Assemblage Development: Suction Sampling
3.2. Comparison of Eelgrass Epifaunal Restored and Natural Communities
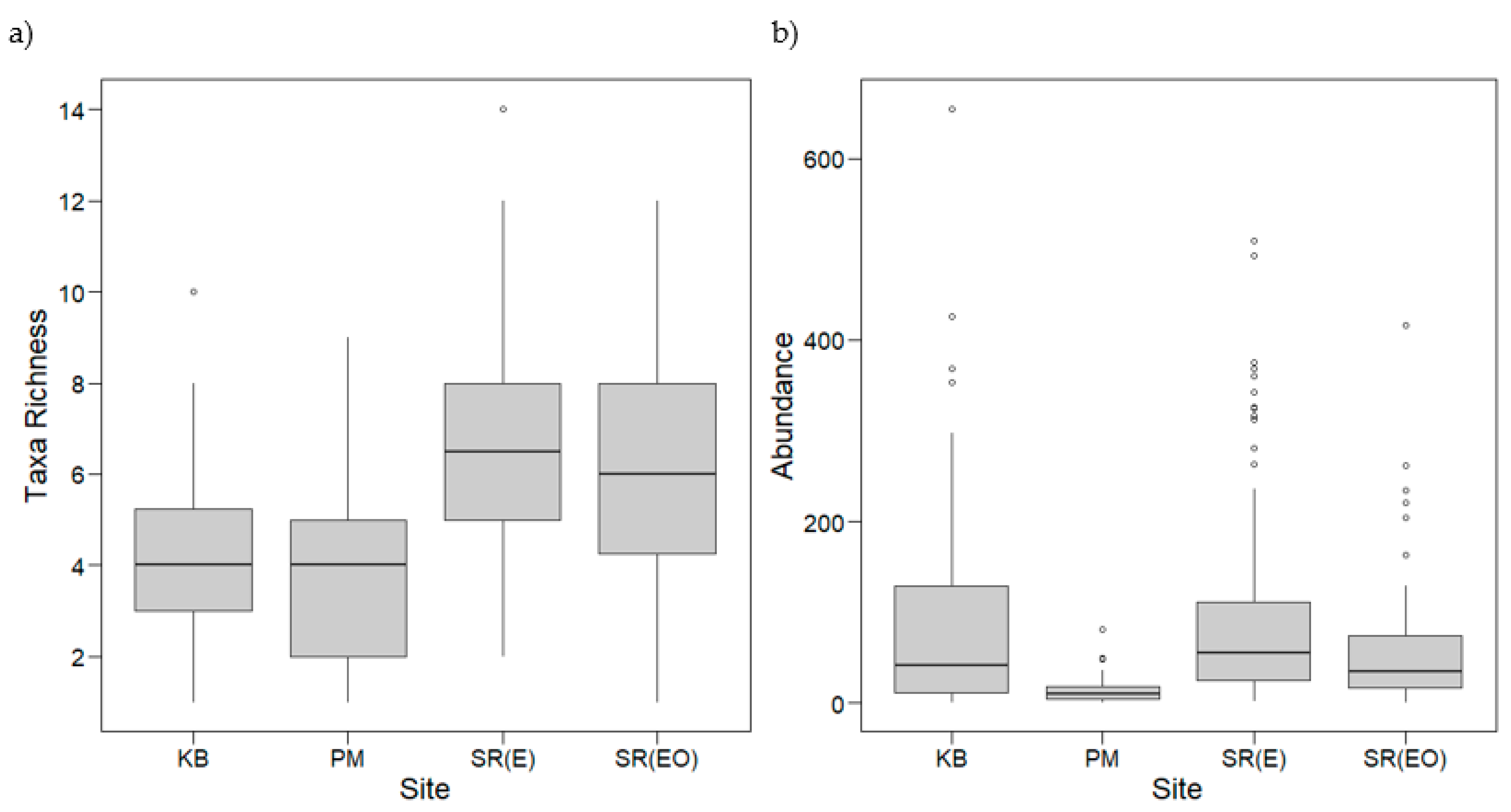
3.2.1. Taxa Richness and Abundance: Shoot Collections
3.2.2. Community Assemblage of Restored and Natural Eelgrass: Shoot Collections
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Treatment | |
|---|---|
| Taxa | SR |
| Fall 2011 | |
| Annelida 2 (Ann2) | 0.3 |
| Corophiidae (Coro) | 1.2 |
| Cumacea (Cuma) | 144.5 |
| Gammarus daiberi (GADA) (g, i) | 73.0 |
| Gastropoda 3 (Gas3) | 0.7 |
| Gemma gemma (GEGE) | 0.7 |
| Grandidierella japonica (GRJA) (g, i) | 11.5 |
| Isopoda (Isop) (I) | 0.7 |
| Potamocorbula amurensis (POAM) (i) | 0.8 |
| Winter 2012 | |
| Annelida 1 (Ann1) | 2.0 |
| Annelida 2 (Ann2) | 22.8 |
| Corophiidae (Coro) | 2.7 |
| Cumacea (Cuma) | 312.8 |
| Gammarus daiberi (GADA) (g, i) | 41.3 |
| Spring 2012 | |
| Annelida 1 (Ann1) | 48.8 |
| Annelida 2 (Ann2) | 13.0 |
| Caprella sp. (CAsp) (c) | 2.2 |
| Corophiidae (Coro) | 39.7 |
| Cumacea (Cuma) | 45.8 |
| Gammarus daiberi (GADA) (g, i) | 70.0 |
| Grandidierella japonica (GRJA) (g, i) | 1.0 |
| Ostracoda (Ostr) | 0.7 |
| Paradexamine sp. (PAsp1) (g) | 0.2 |
| Summer 2012 | |
| Annelida 1 (Ann1) | 0.3 |
| Corophiidae (Coro) | 2.0 |
| Cumacea (Cuma) | 0.2 |
| Gammarus daiberi (GADA) (g, i) | 2.5 |
| Grandidierella japonica (GRJA) (g, i) | 0.2 |
| Paradexamine sp. (PAsp1) (g) | 0.2 |
| Taxa | SREO (E) | SREO (O) | SR E | SR O | SR C |
|---|---|---|---|---|---|
| Summer 2013 | |||||
| Ampithoe valida (AMVA) (g, i) | 0 | 0.7 | 0 | 6.7 | 0 |
| Annelida 1 (Ann1) | 5.0 | 7.7 | 32.5 | 41.3 | 15.3 |
| Annelida 2 (Ann2) | 3.7 | 9.7 | 12.8 | 18.0 | 7.0 |
| Caprella sp. (CAsp) (c) | 0.3 | 0 | 0 | 0.2 | 0.5 |
| Corophiidae (Coro) | 72.0 | 145.5 | 247.5 | 156.3 | 10.7 |
| Cumacea (Cuma) | 14.5 | 4.3 | 54.5 | 5.3 | 34.8 |
| Gammarus daiberi (GADA) (g, i) | 9.3 | 4.8 | 38.3 | 9.8 | 30.8 |
| Gastropoda 3 (Gas3) | 0 | 0 | 4.8 | 0 | 0.3 |
| Grandidierella japonica (GRJA) (g, i) | 66.7 | 47.8 | 31.3 | 78.0 | 70.0 |
| Isopoda (Isop) (I) | 0.2 | 0 | 0 | 0.7 | 0.2 |
| Ostracoda (Ostr) | 0.2 | 0 | 0.2 | 0 | 0.2 |
| Paradexamine sp. (PAsp1) (g) | 0.3 | 8.2 | 0 | 6.2 | 0 |
| Potamocorbula amurensis (POAM) (i) | 0 | 0 | 0.2 | 0 | 0 |
| Fall 2013 | |||||
| Ampithoe valida (AMVA) (g, i) | 3.2 | 0 | 11.7 | 0 | 0 |
| Annelida 1 (Ann1) | 14.3 | 3.5 | 27.8 | 110.3 | 11.3 |
| Annelida 2 (Ann2) | 10.0 | 1.3 | 6.2 | 6.7 | 1.7 |
| Caprella sp. (CAsp) (c) | 3.3 | 0.2 | 0 | 0 | 0 |
| Corophiidae (Coro) | 11.0 | 11.0 | 0.3 | 0.3 | 0 |
| Cumacea (Cuma) | 2.8 | 5.0 | 41.7 | 4.7 | 71.7 |
| Gammarus daiberi (GADA) (g, i) | 0.7 | 0 | 27.3 | 1.3 | 65.8 |
| Gastropoda 1 (Gas1) | 0 | 0.2 | 0 | 0 | 0 |
| Gastropoda 3 (Gas3) | 0 | 0 | 16.7 | 3.7 | 3.3 |
| Gemma gemma (GEGE) (i) | 0 | 0 | 0 | 0.2 | 0 |
| Grandidierella japonica (GRJA) (g, i) | 59.3 | 36.7 | 14.5 | 24.2 | 12.0 |
| Isopoda (Isop) (i) | 0 | 0 | 0.2 | 0.7 | 0 |
| Ostracoda (Ostr) | 0.2 | 0 | 0 | 0.2 | 0.3 |
| Paradexamine sp. (PAsp1) (g) | 0.5 | 0.2 | 0 | 0 | 0 |
| Winter 2014 | |||||
| Amphipoda (Amph) | 0 | 0 | 0 | 0.8 | 0 |
| Ampithoe valida (AMVA) (g, i) | 5.8 | 2.5 | 0 | 0 | 2.5 |
| Annelida 1 (Ann1) | 13.8 | 17.7 | 7.5 | 47.6 | 4.0 |
| Annelida 2 (Ann2) | 42.8 | 67.2 | 2.5 | 21.8 | 22.5 |
| Caprella sp. (CAsp) (c) | 19.2 | 0.5 | 10.7 | 0 | 0 |
| Corophiidae (Coro) | 11.2 | 20.7 | 3.7 | 35.0 | 1.3 |
| Cumacea (Cuma) | 18.0 | 8.7 | 56.8 | 17.2 | 56.2 |
| Gammarus daiberi (GADA) (g, i) | 5.3 | 0.5 | 25.0 | 2.8 | 28.5 |
| Gastropoda 1 (Gas1) | 0 | 0.3 | 0 | 0.8 | 0 |
| Gastropoda 3 (Gas3) | 8.5 | 6.8 | 13.3 | 11.6 | 0.2 |
| Gemma gemma (GEGE) (i) | 0.3 | 0 | 0 | 0.4 | 0 |
| Grandidierella japonica (GRJA) (g, i) | 6.2 | 10.3 | 6.0 | 30.2 | 4.0 |
| Isopoda (Isop) (I) | 0 | 0 | 0.3 | 1.2 | 0 |
| Ostracoda (Ostr) | 0.3 | 0 | 0.7 | 0.2 | 0 |
| Spring 2014 | |||||
| Americhelidium pectinatum (AMPE) (g, n) | 0.3 | 2.3 | 1.7 | 0 | 13.5 |
| Ampelisca abdita (AMAB) (g, i) | 8.8 | 5.5 | 8.8 | 0 | 32.8 |
| Amphipoda (Amph) | 0 | 0 | 0.5 | 0 | 0.5 |
| Ampithoe valida (AMVA) (g, i) | 0 | 0 | 1.3 | 0.8 | 0.2 |
| Annelida 1 (Ann1) | 32.2 | 4.2 | 8.5 | 6.5 | 31.8 |
| Annelida 2 (Ann2) | 3.3 | 2.7 | 3.2 | 17.3 | 8.0 |
| Brachyura (Brac) | 0 | 0 | 0 | 0.2 | 0 |
| Caprella sp. (CAsp) (c) | 0.2 | 0 | 1.8 | 0.7 | 5.3 |
| Corophiidae (Coro) | 23.5 | 11.5 | 7.2 | 53.8 | 54.3 |
| Cumacea (Cuma) | 5.7 | 10.8 | 18.0 | 5.5 | 56.8 |
| Gammarus daiberi (GADA) (g, i) | 0 | 0.3 | 0 | 7.8 | 0 |
| Gastropoda 1 (Gas1) | 0 | 0 | 0 | 0.3 | 0 |
| Gastropoda 3 (Gas3) | 6.7 | 1.2 | 0.8 | 0 | 2.7 |
| Gemma gemma (GEGE) (i) | 0 | 0 | 0.2 | 0.2 | 0 |
| Grandidierella japonica (GRJA) (g, i) | 47.5 | 68.2 | 15.7 | 27.0 | 10.8 |
| Ostracoda (Ostr) | 2.0 | 0.5 | 1.2 | 0.3 | 11.2 |
| Paradexamine sp. (PAsp1) (g) | 1.2 | 0.3 | 14.7 | 6.0 | 63.8 |
| Summer 2014 | |||||
| Americhelidium pectinatum (AMPE) (g, n) | 0 | 0.3 | 15.3 | 16.0 | 7.7 |
| Ampelisca abdita (AMAB) (g, i) | 0.7 | 0.3 | 57.7 | 1.7 | 99.0 |
| Ampithoe valida (AMVA) (g, i) | 0 | 0.3 | 3.0 | 0.3 | 0 |
| Annelida 1 (Ann1) | 0.7 | 3.7 | 12.0 | 7.7 | 10.7 |
| Annelida 2 (Ann2) | 0.3 | 16.3 | 15.7 | 12.3 | 0.3 |
| Caprella sp. (CAsp) (c) | 0 | 0 | 0 | 0 | 0.7 |
| Corophiidae (Coro) | 2.7 | 24.3 | 28.3 | 92.3 | 29.0 |
| Cumacea (Cuma) | 0 | 0 | 12.7 | 0.3 | 20.7 |
| Gammarus daiberi (GADA) (g, i) | 0.3 | 0.3 | 1.3 | 0 | 0 |
| Gastropoda 1 (Gas1) | 0 | 1.3 | 0 | 0.3 | 0 |
| Gastropoda 3 (Gas3) | 0 | 3.3 | 3.7 | 5.7 | 0 |
| Gemma gemma (GEGE) (i) | 0 | 0.3 | 0 | 0 | 0 |
| Grandidierella japonica (GRJA) (g, i) | 1.3 | 18.0 | 25.0 | 6.3 | 43.7 |
| Ostracoda (Ostr) | 0 | 4.0 | 2.7 | 14.3 | 0 |
| Paradexamine sp. (PAsp1) (g) | 4.7 | 13.7 | 0.3 | 4.0 | 0 |
| Potamocorbula amurensis (POAM) (i) | 0 | 0 | 1.3 | 0 | 0 |
| Summer 2015 | |||||
| Americhelidium pectinatum (AMPE) (g, n) | - | 2.0 | 36.0 | 22.3 | 52.3 |
| Amphipoda (Amph) | - | 0.7 | 0.3 | 3.0 | 0 |
| Annelida 1 (Ann1) | - | 4.3 | 0.7 | 0 | 38.7 |
| Annelida 2 (Ann2) | - | 20.7 | 5.0 | 9.3 | 44.3 |
| Caprella sp. (CAsp) (c) | - | 0.3 | 0 | 0 | 0 |
| Cerapus sp. (CEsp) (g) | - | 0.7 | 0 | 8.3 | 0.3 |
| Corophiidae (Coro) | - | 9.3 | 1.3 | 5.3 | 0.3 |
| Cumacea (Cuma) | - | 1.7 | 5.7 | 5.3 | 67.7 |
| Daphnia sp. (DAsp) | - | 2.7 | 0 | 0 | 0 |
| Gastropoda 3 (Gas3) | - | 0.7 | 2.7 | 3.0 | 1.3 |
| Grandidierella japonica (GRJA) (g, i) | - | 6.7 | 1.7 | 6.7 | 54.7 |
| Munna sp. (Musp) | - | 16.0 | 1.0 | 2.0 | 0 |
| Ostracoda (Ostr) | - | 1.7 | 2.0 | 3.7 | 0.7 |
| Paradexamine sp. (PAsp1) (g) | - | 7.0 | 3.3 | 3.0 | 0 |
| Paranthura sp. (PAsp2) (i) | - | 2.3 | 1.3 | 2.0 | 4.0 |
| Polyplacophora (Poly) | - | 0 | 0.3 | 0 | 0 |
| KB | PM | SR | SR | |
|---|---|---|---|---|
| Taxa | Natural | Natural | Eelgrass | Eelgrass + Oyster |
| Year 1 | ||||
| Summer 2013 | ||||
| Ampelisca abdita (AMAB) (g, i) | - | - | 7.3 | 0.2 |
| Ampithoe valida (AMVA) (g, i) | - | - | 11.7 | 17.6 |
| Annelida 1 (Ann1) | - | - | 0.8 | 3.9 |
| Annelida 2 (Ann2) | - | - | 1 | 2.3 |
| Brachyura (Brac) | - | - | 0.1 | 0 |
| Caprella sp. (CAsp) (c) | - | - | 1.3 | 1.3 |
| Cirripedia (Cirr) | - | - | 0.5 | 0.3 |
| Corophiidae (Coro) (g) | - | - | 218.7 | 129.3 |
| Cumacea (Cuma) | - | - | 0.1 | 0.1 |
| Gammarus daiberi (GADA) (g, i) | - | - | 0.3 | 1.2 |
| Gastropoda 2 (Gas2) | - | - | 1.1 | 0.1 |
| Gastropoda 3 (Gas3) | - | - | <0.1 | 0.1 |
| Grandidierella japonica (GRJA) (g, i) | - | - | 0.3 | 1.8 |
| Isopoda (Isop) (I) | - | - | <0.1 | <0.1 |
| Ostracoda (Ostr) | - | - | 0 | 0.3 |
| Paradexamine sp. (PAsp1) (g) | - | - | 14 | 4.1 |
| Phyllaplysia taylori (PHTA) (n) | - | - | 0 | 0.1 |
| Amphipoda (Amph) | - | 0.7 | 1.1 | 4.1 |
| Ampithoe valida (AMVA) (g, i) | - | 12.3 | 19.4 | 25.9 |
| Annelida 1 (Ann1) | - | 0 | 0 | 0.2 |
| Annelida 2 (Ann2) | - | 0.7 | 0.2 | 1.6 |
| Caprella californica (CACA) (c, n) | - | 0 | <0.1 | 0.2 |
| Caprella drepanochir (CADR) (c, i) | - | 1.7 | 0 | 0 |
| Caprella sp. (CAsp) (c) | - | 0 | 46.3 | 39.4 |
| Cirripedia (Cirr) | - | 0 | 13.7 | 14.3 |
| Corophiidae (Coro) (g) | - | 0 | 24.2 | 12.9 |
| Cumacea (Cuma) | - | 0 | 0 | 0.1 |
| Gammarus daiberi (GADA) (g, i) | - | 0 | 0 | 0.2 |
| Grandidierella japonica (GRJA) (g, i) | - | 0 | 2.4 | 11.4 |
| Jassa slatteryi (JASL) (g, c) | - | 9.3 | 0 | 0 |
| Ostracoda (Ostr) | - | 1 | 0 | 0.1 |
| Paradexamine sp. (PAsp1) (g) | - | 0 | 2.2 | 8.9 |
| Pentidotea resecata (PERE) (I, n) | - | 1 | 0 | 0 |
| Phyllaplysia taylori (PHTA) (n) | - | 3 | 0 | 0 |
| Synidotea laticauda (SYLA) (I, i) | - | 0.3 | 0 | 0 |
| Spring 2014 | ||||
| Amphipoda (Amph) | 1.6 | 0 | 0.1 | 0 |
| Ampithoe valida (AMVA) (g, i) | 0.9 | 0 | 2.1 | 0.7 |
| Annelida 1 (Ann1) | 0.2 | 0.8 | 0.2 | 0.8 |
| Annelida 2 (Ann2) | 0.5 | 3.5 | 0.6 | 0.7 |
| Brachyura (Brac) | 0.2 | 0.2 | <0.1 | 0 |
| Caprella sp. (CAsp) (c) | 128.5 | 0.4 | 48 | 2.5 |
| Cirripedia (Cirr) | 0 | 0 | 9.5 | 0.9 |
| Corophiidae (Coro) (g) | 0.4 | 0.5 | 90.9 | 17.3 |
| Gammarus daiberi (GADA) (g, i) | 0 | 0.2 | 0.6 | 2.2 |
| Gastropoda 1 (Gas1) | 0 | 0 | 0.1 | 0 |
| Gastropoda 3 (Gas3) | 0.3 | 0.1 | 0.1 | 0.6 |
| Grandidierella japonica (GRJA) (g, i) | 31.8 | 1.8 | 2.7 | 2.3 |
| Ostracoda (Ostr) | 0 | 0 | 0.2 | 0.1 |
| Paradexamine sp. (PAsp1) (g) | 0.3 | 1.7 | 7.3 | 6.1 |
| Pentidotea resecata (PERE) (I, n) | 1 | 11.5 | 0 | 0 |
| Phyllaplysia taylori (PHTA) (n) | 0 | 1.3 | 0 | 0 |
| Year 2 | ||||
| Summer 2014 | ||||
| Ampelisca abdita (AMAB) (g, i) | - | - | 0.1 | 0.1 |
| Ampithoe valida (AMVA) (g, i) | - | - | 33.1 | 33.6 |
| Annelida 1 (Ann1) | - | - | 0.3 | 6.3 |
| Annelida 2 (Ann2) | - | - | 3.8 | 2.5 |
| Bivalvia (Biva) | - | - | 0.3 | 0.2 |
| Caprella sp. (CAsp) (c) | - | - | 0.3 | 1.5 |
| Cirripedia (Cirr) | - | - | 1.8 | 1.3 |
| Corophiidae (Coro) (g) | - | - | 1049.5 | 471.9 |
| Gastropoda 1 (Gas1) | - | - | 0 | 0.2 |
| Gastropoda 3 (Gas3) | - | - | 0 | 0.5 |
| Gemma gemma (GEGE) (i) | - | - | 0.2 | 0.8 |
| Grandidierella japonica (GRJA) (g, i) | - | - | 2.1 | 0.1 |
| Isopoda (Isop) (I) | - | - | 0 | 0.1 |
| Jassa slatteryi (JASL) (g, c) | - | - | 75.1 | 8.8 |
| Jassa sp. (JAsp) (g) | - | - | 0.7 | 0 |
| Ostracoda (Ostr) | - | - | <0.1 | 1.6 |
| Paradexamine sp. (PAsp1) (g) | - | - | 0.9 | 1.9 |
| Phyllaplysia taylori (PHTA) (n) | - | - | 0.1 | 0 |
| Platyhelminthes (Plat) | - | - | 0 | <0.1 |
| Potamocorbula amurensis (POAM) (i) | - | - | 0 | <0.1 |
| Siliqua patula (SIPA) (n) | - | - | 0 | 0.8 |
| Tanaidacea (Tana) | - | - | <0.1 | 0.5 |
| Fall 2014 | ||||
| Ampelisca abdita (AMAB) (g, i) | 0 | 0 | 0.1 | 0.1 |
| Amphipoda (Amph) | 0 | 0 | 7.5 | 1 |
| Ampithoe valida (AMVA) (g, i) | 2.2 | 4.4 | 5.5 | 2.9 |
| Annelida 1 (Ann1) | 0 | 0 | 1.5 | 2.1 |
| Annelida 2 (Ann2) | 1 | 1.1 | 1.5 | 2.9 |
| Bivalvia (Biva) | 0 | 0 | <0.1 | 0.1 |
| Caprella californica (CACA) (c, n) | 0 | 0 | 0.3 | 0.2 |
| Caprella sp. (CAsp) (c) | 0 | 0 | 8.6 | 1.1 |
| Cirripedia (Cirr) | 0 | 0 | <0.1 | 0.5 |
| Corophiidae (Coro) (g) | 0.5 | 1.6 | 1 | 6.4 |
| Cumacea (Cuma) | 0 | 0 | 0 | <0.1 |
| Gammarus daiberi (GADA) (g, i) | 0 | 0 | 0 | 0.1 |
| Gastropoda 3 (Gas3) | 0 | 0 | 0.1 | 0 |
| Gemma gemma (GEGE) (i) | 0 | 0 | 0 | <0.1 |
| Gnorimosphaeroma oregonensis (GNOR) (I, n) | 0 | 0 | <0.1 | 0.1 |
| Grandidierella japonica (GRJA) (g, i) | 0 | 0 | 0 | 0.1 |
| Isopoda (Isop) (I) | 0 | 0 | 0 | 0 |
| Jassa slatteryi (JASL) (g, c) | 0 | 0.1 | 56.8 | 5.9 |
| Jassa sp. (JAsp) (g) | 0 | 0 | 0.1 | 0.1 |
| Ostracoda (Ostr) | 0 | 0.1 | 1 | 5 |
| Paradexamine sp. (PAsp1) (g) | 0 | 0 | 23.3 | 38 |
| Paranthura sp. (PAsp2) (i) | 0 | 0 | 0.5 | 0.5 |
| Pentidotea resecata (PERE) (I, n) | 0 | 0 | 0 | <0.1 |
| Phyllaplysia taylori (PHTA) (n) | 0 | 0.7 | 0 | 0 |
| Platyhelminthes (Plat) | 0 | 0 | <0.1 | 0 |
| Stenothoe valida (STVA) (g, i) | 0 | 0 | 20.9 | <0.1 |
| Synidotea laticauda (SYLA) (i, i) | 0 | 0 | 0.3 | 0.1 |
| Tanaidacea (Tana) | 0.2 | 1 | 0.2 | 0.2 |
| Allorchestes angusta (ALAN) (g, n) | 0 | 0 | 0.1 | 0.1 |
| Amphipoda (Amph) | 6.4 | 0.1 | 3.4 | 4.9 |
| Ampithoe valida (AMVA) (g, i) | 0.1 | 1.2 | 6.1 | 3.9 |
| Annelida 1 (Ann1) | 0.4 | 0 | 3.2 | 50.4 |
| Annelida 2 (Ann2) | 0.1 | 0.6 | 2.8 | 3.9 |
| Bivalvia (Biva) | 0 | 0 | 0.1 | 0.3 |
| Caprella californica (CACA) (c, n) | 1.6 | 3.2 | 2.7 | 0.7 |
| Caprella drepanochir (CADR) (c, i) | 5.9 | 0 | 0.1 | 0 |
| Caprella sp. (CAsp) (c) | 125.1 | 4.4 | 6.3 | 0.3 |
| Cirripedia (Cirr) | 0 | 0 | 0.4 | 0 |
| Corophiidae (Coro) (g) | 0 | 0 | 11.5 | 8.8 |
| Cumacea (Cuma) | 31.5 | 0 | 0.1 | 0.1 |
| Gammarus daiberi (GADA) (g, i) | 0 | 0 | 0 | 0.1 |
| Gastropoda 1 (Gas1) | 0 | 0 | 0.3 | 0 |
| Gastropoda 3 (Gas3) | 0 | 0 | 0.9 | 2.3 |
| Gnorimosphaeroma oregonensis (GNOR) (I, n) | 0 | 0 | 0.1 | 0 |
| Jassa slatteryi (JASL) (g, c) | 1.1 | 0.3 | 1 | 0.4 |
| Ostracoda (Ostr) | 0 | 0 | 14.5 | 23.4 |
| Paradexamine sp. (PAsp1) (g) | 0.3 | 0 | 0.2 | 0.5 |
| Paranthura sp. (PAsp2) (i) | 0 | 0 | 0 | 0.1 |
| Pentidotea resecata (PERE) (I, n) | 0.1 | 0 | 0 | 0 |
| Phyllaplysia taylori (PHTA) (n) | 0 | 1.1 | 0.2 | 0 |
| Platyhelminthes (Plat) | 0 | 2.2 | <0.1 | 0.9 |
| Stenothoe valida (STVA) (g, i) | 0 | 0 | 0.3 | 0.1 |
| Tanaidacea (Tana) | 0 | 0 | 0.3 | 1 |
| Year 3 | ||||
| Summer 2015 | ||||
| Allorchestes angusta (ALAN) (g, n) | 0 | 0 | 0 | 0.1 |
| Amphipoda (Amph) | 6.2 | 1.3 | 1.1 | 1.3 |
| Ampithoe lacertosa (AMLA) (g, n) | 0.1 | 0 | 0.1 | 0.1 |
| Ampithoe valida (AMVA) (g, i) | 0.6 | 0.4 | 3.5 | 3.1 |
| Annelida 1 (Ann1) | 0.2 | 0 | 0.4 | 6.2 |
| Annelida 2 (Ann2) | 3.7 | 3.1 | 8.2 | 10.8 |
| Caprella californica (CACA) (c, n) | 0.5 | 0.8 | 1.6 | 1.6 |
| Caprella drepanochir (CADR) (c, i) | 1.3 | 0 | 0 | 0.1 |
| Caprella sp. (CAsp) (c) | 3.1 | 0 | 0.1 | 0.8 |
| Cirripedia (Cirr) | 0 | 0 | 0.1 | 0 |
| Corophiidae (Coro) (g) | 1.5 | 1.4 | 5.8 | 2.9 |
| Cumacea (Cuma) | 0 | 0 | 0.5 | 0.1 |
| Gastropoda 3 (Gas3) | 0.5 | 0 | 2 | 2.1 |
| Gemma gemma (GEGE) (i) | 0 | 0.1 | 0 | 0 |
| Grandidierella japonica (GRJA) (g, i) | 0 | 0 | 0 | 0.2 |
| Jassa slatteryi (JASL) (g, c) | 13.4 | 5.2 | 0.1 | 0 |
| Ostracoda (Ostr) | 0.2 | 0.5 | 3.1 | 10.6 |
| Paradexamine sp. (PAsp1) (g) | 1.5 | 0.7 | 1.6 | 0.7 |
| Paranthura sp. (PAsp2) (i) | 0 | 0.2 | 1.5 | 0.7 |
| Pentidotea resecata (PERE) (I, n) | 0.2 | 1.4 | 0 | 0 |
| Potamocorbula amurensis (POAM) (i) | 0 | 0 | 0.1 | 0 |
| Synidotea laticauda (SYLA) (I, i) | 0 | 0 | 0.1 | 0 |
| Tanaidacea (Tana) | 0.1 | 0 | 0 | 0 |
| Factor | Class 1 | Pr(>F) | r2 |
|---|---|---|---|
| Years 0 and 1 2 | |||
| Treatment | P, E, EO(E), O, EO(O), C | <0.001 | 0.25 |
| Season | Fall, Spring, Summer, Winter | <0.001 | 0.15 |
| Treatment × Season | <0.001 | 0.23 | |
| Years 0, 1, 2, and 3 3 | |||
| Treatment | P, E, EO(E), O, EO(O), C | <0.001 | 0.21 |
| Season | Fall, Spring, Summer, Winter | <0.001 | 0.11 |
| Treatment × Season | <0.001 | 0.20 | |
| Years 0, 1, 2, and 3 4 | |||
| Treatment | P, E, EO(E), O, EO(O), C | <0.001 | 0.26 |
| Year | 0, 1, 2, and 3 | <0.001 | 0.34 |
| Treatment × Year | <0.001 | 0.16 | |
| Factor | Class 1 | Pr(>F) | r2 |
|---|---|---|---|
| Treatment | E, EO, N | <0.001 | 0.07 |
| Source Site | SR(E), SR(EO), PM, KB | <0.001 | 0.10 |
| Year | 1,2,3 | <0.001 | 0.10 |
| Treatment × Year | <0.001 | 0.08 | |
| Source Site × Year | <0.001 | 0.10 |



References
- Bell, S.E.; McCoy, E.; Mushinsky, H. Habitat structure: The physical arrangement of objects in space. In Chapman & Hall; Springer: Dordrecht, The Netherlands, 1991. [Google Scholar]
- Byrne, L. Habitat Structure: A fundamental concept and framework for urban soil ecology. Urban Ecosyst. 2007, 10, 255–274. [Google Scholar] [CrossRef]
- Heck, K.L.; Wetstone, G.S. Habitat complexity and invertebrate species richness and abundance in tropical seagrass meadows. J. Biogeogr. 1977, 4, 135–142. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat complexity: Approaches and future directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- MacArthur, R.H. Patterns of species diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliot, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Guitierrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Schmidt, A.L.; Coll, M.; Romanuk, T.N.; Lotze, H.K. Ecosystem structure and services in eelgrass Zostera marina and rockweed Ascophyllum nodosum habitats. Mar. Ecol. Prog. Ser. 2011, 437, 51–68. [Google Scholar] [CrossRef] [Green Version]
- Rozas, L.P.; Minello, T.J. Nekton use of salt marsh, seagrass, and non-vegetated habitats in a south Texas (USA) estuary. Bull. Mar. Sci. 1998, 63, 481–501. [Google Scholar]
- Glancy, T.P.; Frazer, T.K.; Cichra, C.E.; Lindberg, W.J. Comparative patterns of occupancy by decapod crustaceans in seagrass, oyster, and marsh-edge habitats in a Northeast Gulf of Mexico estuary. Estuaries 2003, 26, 1291–1301. [Google Scholar] [CrossRef]
- Hosack, G.R.; Dumbauld, B.R.; Ruesink, J.L.; Armstrong, D.A. Habitat associations of estuarine species: Comparisons of intertidal mudflat, seagrass (Zostera marina), and oyster (Crassostrea gigas) habitats. Estuaries Coasts 2006, 29, 1150–1160. [Google Scholar] [CrossRef]
- Orth, R.T.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, A.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, M.S.; Fisher, J.S.; Ziemen, J.C.; Thayer, G.W. Influence of the seagrass (Zostera marina L.) on current flow. Estuarine Coast. Shelf Sci. 1982, 15, 351–364. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Marcomini, A. Sedimentation rates and erosion processes in the Lagoon of Venice. Environ. Int. 2005, 33, 274. [Google Scholar]
- Bouma, T.J.; van Duren, L.A.; Temmerman, S.; Claverie, T.; Blano-Garcia, A.; Ysebaert, T.; Herman, P.J. Spatial flow and sedimentation patterns within patches of epibenthic structures: Combining field, flume and modeling experiments. Cont. Shelf Res. 2005, 27, 1020–1045. [Google Scholar] [CrossRef]
- Bos, A.R.; Bouma, T.J.; de Kort, G.L.; van Katwijk, M.M. Ecosystem engineering by annual intertidal seagrass beds: Sediment accretion and modification. Estuarine Coast. Shelf Sci. 2007, 74, 344–348. [Google Scholar] [CrossRef]
- Dame, R.J.; Spurrier, J.; Wolaver, T. Carbon, nitrogen and phosphorous processing by an oyster reef. Mar. Ecol. Prog. Ser. 1999, 54, 249–256. [Google Scholar] [CrossRef]
- Polson, M.P.; Zacherl, D.C. Geographic distribution and intertidal population status for the Olympia oyster, Ostrea lurida Carpenter 1864, from Alaska to Baja. J. Shellfish Res. 2009, 28, 69–77. [Google Scholar] [CrossRef]
- Schulte, D.M.; Burke, R.P.; Lipcius, R.N. Unprecedented Restoration of a Native Oyster Metapopulation. Science 2009, 325, 1124–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, C. The future of seagrass meadows. Environ. Conserv. 2002, 29, 192–206. [Google Scholar] [CrossRef] [Green Version]
- Erftemeijer, P.; Lewis, R., III. Environmental impacts of dredging on seagrasses: A review. Mar. Pollut. Bull. 2006, 52, 1553–1572. [Google Scholar] [CrossRef]
- California State Coastal Conservancy (SCC). San Francisco Bay Subtidal Habitat Goals Report: Conservation Planning for the Submerged Areas of the Bay; California State Coastal Conservancy (SCC): Oakland, CA, USA, 2010.
- Kairis, P.A.; Rybczyk, J.M. Sea level rise and eelgrass (Zostera marina) production: A spatially explicit relative elevation model for Padilla Bay, WA. Ecol. Model. 2010, 221, 1005–1016. [Google Scholar] [CrossRef]
- California Department of Fish and Game. California Wildlife Action Plan Report: Central Valley and Bay-Delta Region; 2007. Available online: http://www.dfg.ca.gov/wildlife/wap/report.html (accessed on 3 November 2011).
- San Francisco Estuary Partnership. The State of the San Francisco Bay; San Francisco Estuary Partnership: Oakland, CA, USA, 2011. [Google Scholar]
- California Department of Fish and Game. 2011 California legislative fisheries forum. In Dept of Fish and Game Annual Marine Fisheries Report; California Department of Fish and Game, Marine Region (Region 7): Monterey, CA, USA, 2011. [Google Scholar]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Metropolitan Transportation Commission—Association of Bay Area Governments, Bay Area Census. 2011. Available online: http://www.bayareacensus.ca.gov/ (accessed on 3 November 2011).
- Heck, K.L.; Orth, R.J. Structural components of eelgrass (Zostera marina) meadows in the lower Chesapeake Bay—decapod crustacean. Estuaries Coasts 1980, 3, 289–295. [Google Scholar] [CrossRef]
- Pauley, G.B.; Armstrong, D.A.; Heun, T.W. Species profiles: Life histories and environmental requirements of coastal fishes and invertebrates (Pacific Northwest)—Dungeness crab. US Fish and Wildlife Service. Bioligical Rep. 1986, 82, 20. [Google Scholar]
- Gunderson, D. Patterns of estuarine use by juvenile English sole (Parophrys vetulus) and Dungeness crab (Cancer magister). Estuaries 1990, 13, 59–71. [Google Scholar] [CrossRef]
- Armstrong, D.; Rooper, C.; Gunderson, D. Estuarine production of juvenile Dungeness crab (Cancer magister) and contribution to the Oregon-Washington coastal fishery. Estuaries 2003, 26, 1174–1188. [Google Scholar] [CrossRef]
- Holsman, K.K.; Armstrong, D.A.; Beauchamp, D.A.; Ruesink, J.L. The necessity for intertidal foraging by estuarine populations of subadult Dungeness crab, Cancer magister: Evidence from a Bioenergetics Model. Estuaries 2003, 26, 1155–1173. [Google Scholar] [CrossRef]
- Boyer, K.E.; Wyllie-Echeverria, S. Eelgrass conservation and Restoration in San Francisco Bay: Opportunities and Constraints. San Francisco Bay Subtidal Habitat Goals Project. Appendix 8-1. 2010. Available online: www.sfbaysubtidal.org/report.html (accessed on 4 March 2016).
- Carr, L.A.; Boyer, K.E.; Brooks, A.J. Spatial patterns of epifaunal communities in San Francisco Bay eelgrass (Zostera marina) beds. Mar. Ecol. 2011, 32, 88–103. [Google Scholar] [CrossRef]
- Duffy, J.E.; Reynolds, P.E.; Bostrom, C.; Coyer, A.J.; Cusson, M.; Donadi, S.; Douglass, J.G.; Eklof, J.S.; AEngelen, H.; Eriksson, B.K.; et al. Biodiversity mediates top-down control in eelgrass ecosystems; a global comparative-experimental approach. Ecol. Lett. 2015, 18, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.F.; Blanchet, G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; et al. Vegan: Community Ecology Package. R Package Version 2.0–10. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 24 February 2014).
- Reynolds, L.K.; Carr, L.A.; Boyer, K.E. A non-native amphipod consumes eelgrass inflorescences in San Francisco Bay. Mar. Ecol. Prog. Ser. 2012, 451, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Carr, L.A.; Boyer, K.E. Variation at multiple trophic levels mediates a novel seagrass-grazer interaction. Mar. Ecol. Prog. Ser. 2014, 508, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.T.; Boyer, K.E. Grazer functional roles, induced defenses, and indirect interactions: Implications for eelgrass restoration in San Francisco Bay. Diversity 2014, 6, 751–770. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.E.; Orth, R.J.; Lefcheck, J.S. Habitat primarily structures seagrass epifaunal communities: A regional-scale assessment in the Chesapeake Bay. Estuaries Coasts 2021, 44, 442–452. [Google Scholar] [CrossRef]
- Weaver, C.L. The Effects of Sediments and Associated Microbial Communities in Zostera marina Restoration. Master’s Thesis, San Francisco State University, San Francisco, CA, USA, 2017. [Google Scholar]
- Pinnell, C. Invertebrate Response to Eelgrass and Oyster Restoration in San Francisco Estuary. Master’s Thesis, San Francisco State University, San Francisco, CA, USA, 2016. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinnell, C.M.; Ayala, G.S.; Patten, M.V.; Boyer, K.E. Seagrass and Oyster Reef Restoration in Living Shorelines: Effects of Habitat Configuration on Invertebrate Community Assembly. Diversity 2021, 13, 246. https://doi.org/10.3390/d13060246
Pinnell CM, Ayala GS, Patten MV, Boyer KE. Seagrass and Oyster Reef Restoration in Living Shorelines: Effects of Habitat Configuration on Invertebrate Community Assembly. Diversity. 2021; 13(6):246. https://doi.org/10.3390/d13060246
Chicago/Turabian StylePinnell, Cassie M., Geana S. Ayala, Melissa V. Patten, and Katharyn E. Boyer. 2021. "Seagrass and Oyster Reef Restoration in Living Shorelines: Effects of Habitat Configuration on Invertebrate Community Assembly" Diversity 13, no. 6: 246. https://doi.org/10.3390/d13060246